Bot. Bull. Acad. Sin. (1995) 36: 143-153
Chiang Phylogeny of Macrothamnium
Phylogeny and morphological evolution of Macrothamnium M. Fleisch. and related taxa (Bryopsida: Hypnaceae)
Tzen-Yuh Chiang
Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
(Received February 27, 1995; Accepted June 5, 1995)
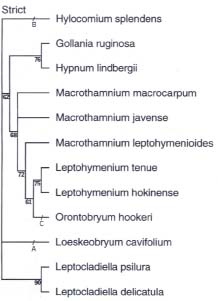
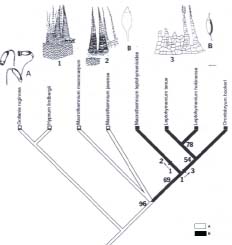
Abstract. Three species of Macrothamnium M. Fleisch., two species of Leptohymenium Schwaegr., two species of Leptocladiella M. Fleisch., and one species of Orontobryum M. Fleisch. were revised. Macrothamnium submacrocarpum Renauld & Cardot and M. longirostre Dix. were synonymized with M. macrocarpum (Reinw. & Hornsch.) M. Fleisch. Macrothamnium hylocomioides M. Fleisch. and Chaetomitriopsis diversifolia Zanten were considered identical to M. javense M. Fleisch. A new combination, Hypnum flagellaris (T. J. Kop. & D. H. Norris) T.Y. Chiang, was proposed and excluded from genus Leptocladiella. The phylogeny of Macrothamnium and the related genera was reconstructed based on ontogenetic transformations of paraphyllia, axillary hairs, central strand, and other morphological characters. Whole ontogenetic transformations, instead of instantaneous stages, were recognized as characters and polarized by outgroup comparison. To test the phylogenetic relationship (familial position), Hypnum lindbergii Mitt. and Gollania ruginosa (Mitt.) Broth. of the Hypnaceae and Hylocomium splendens (Hedw.) B.S.G. and Loeskeobryum cavifolium (Lac.) M. Fleisch. of Hylocomiaceae were chosen as outgroups. The monophyly of the clade of Macrothamnium, Leptohymenium, and Orontobryum was supported, with a bootstrapping value of 96%, and was characterized by sharing regularly pinnate branching pattern, amplified costa, and a lack of foliose pseudoparaphyllia, whereas Macrothamnium appeared to be a paraphyletic group, in which M. leptohymenioides Nog. is more related to Leptohymenium than it is to any other species of Macrothamnium. The three genera are more related to the Hypnaceae than to the Hylocomiaceae. In contrast, Leptocladiella appears to be a genus of Hylocomiaceae. Patterson's tests were applied to the homology of the horn-type paraphyllia of Hylocomium splendens and Loeskeobryum cavifolium, and the foliose-type paraphyllia of Orontobryum hookeri. They passed the similarity and conjunction tests, but failed the congruence test. This suggests that the two types of paraphyllia are homoplasticthat is, they evolved independently rather than being derived from a most recent ancestor.
Keywords: Homology; Macrothamnium; Monophyly; Ontogenetic transformations; Paraphyllia; Patterson's three tests; Phyllodioicous.
defined by weft growth-form. In delimiting the genera of the Hypnaceae, Nishimura et al. (1984) adopted Andrews' and Noguchi's concepts, and classified Macrothamnium, Leptocladiella, and Leptohymenium as Hypnaceae and Orontobryum as Hylocomiaceae by using the presence or absence of paraphyllia to distinguish them.
Buck and Vitt (1986) criticized the sole use of paraphyllia in defining the Hylocomiaceae as illogical. In their so-called `pseudo-cladistic analysis', which was performed by arbitrary and empirical interpretation of morphological characters and taxonomic relationships of pleurocarpous mosses, they characterized the Hylocomiaceae by serrate leaf-margins, amplified costa, and reticulate exostome ornamentation. They classified Macrothamnium, Leptocladiella, Leptohymenium, and Orontobryum as Hylocomiaceae.
Buck and Crum (1990), in an evaluation of familial limits of Thuidiaceae and Leskeaceae, re-emphasized paraphyllia in defining the Hylocomiaceae and transferred genera Hylocomiopsis and Actinothuidium into the Hylocomiaceae. This classification has increased the complexity of the circumscription of the Hylocomiaceae and related families.
Introduction
The genera Macrothamnium, Leptocladiella, Leptohymenium, and Orontobryum are distributed mainly in the montane regions of southeastern Asia, with one exceptionLeptohymenium tenue (Hook.) Schwaegr., which was reported in the New World (Mexico) (Rohrer, 1985b, 1986). According to the fossil record of a Macrothamnium sp. found in Poland, the origin of the taxa can be traced to the Miocene (Miller, 1984).
The close relationship of Macrothamnium, Leptohymenium, and Orontobryum has been interpreted in Rohrer's cladistic study on the Hylocomiaceae (Rohrer, 1985a), based on eighteen morphological characters. He recognized Leptocladiella as a genus more distantly related to the above taxa, although many other bryologists had synonymized it either to Leptohymenium (Andrews, 1954) or to Macrothamnium (Noguchi, 1972a).
The taxonomic position of the taxa is a controversial issue. There has been conjecture about the familial position of Macrothamnium, Leptocladiella, Leptohymenium, and Orontobryum. Rohrer (1985b) recognized twelve genera, including the above four taxa, in the Hylocomiaceae,