Bot. Bull. Acad. Sin. (1995) 36: 169-173
Lin and Kao Gibberellic acid, starch mobilization and NaCl stress
NaCl stress in rice seedlings: starch mobilization and the
influence of gibberellic acid on seedling growth
Chuan Chi Lin and Ching Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received January 6, 1995; Accepted April 12, 1995)
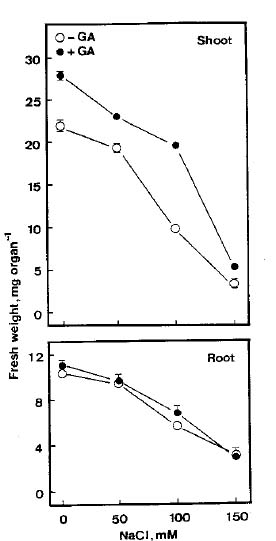
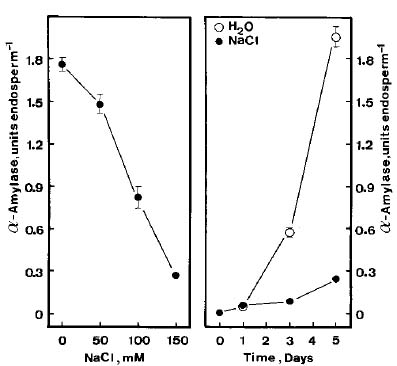
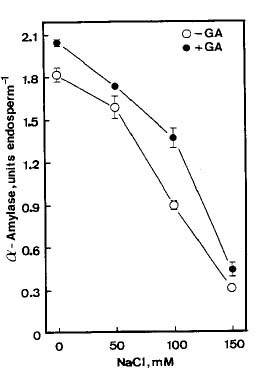
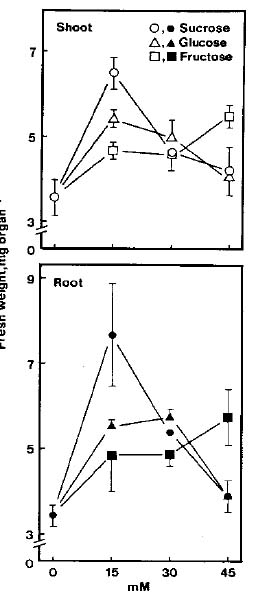
Abstract. The growth of shoots and roots of rice (Oryza sativa L., cv. Taichung Native 1) seedlings was significantly inhibited when the seeds were subjected to NaCl stress. NaCl markedly decreased the mobilization of starch in endosperm. Results also showed that a-amylase activities in endosperm were reduced when rice seeds were germinated in the presence of NaCl. NaCl-inhibition of a-amylase activities was counteracted by gibberellic acid (GA3). GA3 reduced NaCl-inhibition of shoot growth, but not of root growth. Sugars (sucrose, fructose, and glucose) were able to reduce NaCl-induced growth inhibition of shoots and roots. The possible mechanism by which shoot growth and root growth in NaCl media respond differently to GA3 is discussed.
Keywords: a-Amylase; Gibberellic acid; NaCl; Oryza sativa L.; Starch mobilization.
Abbreviation: GA3, gibberellic acid.
Introduction
Mobilization of seed reserves, which occurs during early seed germination, is crucial because it supplies substrates for the proper functioning of different metabolic processes that are essential to growth of the embryonic axis (Mayer and Poljakoff-Mayber, 1975). Rice is a salt-sensitive crop species (Flowers and Yeo, 1981). The mechanism of NaCl inhibition of rice-seedling growth is unclear, but NaCl may inhibit mobilization of seed reserves (Prakash and Prathapasenan, 1988).
Gibberellic acid (GA3) is known to induce the synthesis of a-amylase in embryo-less rice seeds (Palmiano and Juliano, 1972). It is not known whether GA3 can reduce NaCl inhibition of rice-seedling growth, but GA3 has been reported to promote the growth of cotton and some halophytes in saline condition (Agakishiev, 1964; Boucaud and Ungar, 1976a; 1976b; Zhao et al., 1986). Huber et al. (1974) also reported that GA3 counteracted the influence of NaCl on the carbohydrate metabolism in leaves of Pennisetum typhoides. The present investigation was conducted to examine whether NaCl-inhibited growth of rice seedlings is mediated through diminishing mobilization of starch in endosperm, and to determine the influence of GA3 on NaCl-inhibited rice-seedling growth.
Materials and Methods
Rice (Oryza sativa L. cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypochlorite for 15 min and washed thoroughly with distilled water. These seeds were
then germinated for 1 day in petri dishes (20 cm) containing distilled water at 37°C in the dark. Uniformly germinated seeds were selected and transferred to petri dishes (9 cm) containing two sheets of Whatman No. 1 filter paper moistened with 10 ml of distilled water or test solution. Each petri dish contained 20 germinated seeds. Each treatment was performed 4 times. The germinated seeds were allowed to grow at 27°C in darkness, and 3 ml of distilled water or test solution was added to each petri dish on day 3.
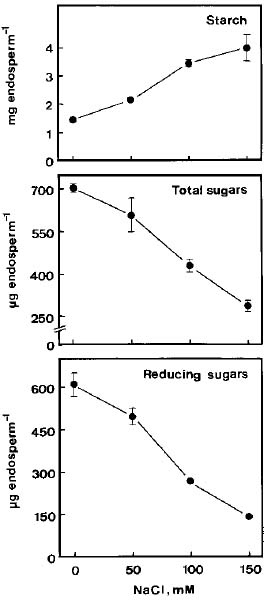
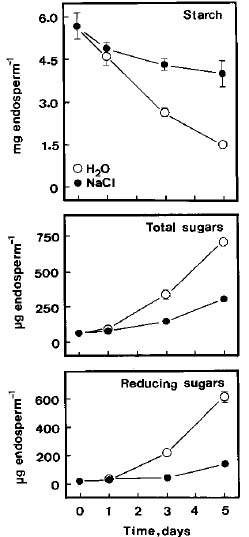
Sugars and starch were extracted from the endosperm twice with hot ethanol (80%). The extract was evaporated to dryness and the residue was dissolved in 2 ml of distilled water. A portion of this extract was used for the estimation of total soluble sugars and reducing sugars using the methods of Yoshida et al. (1972) and Lindsay (1973), respectively. Total soluble sugars and reducing sugars are expressed as mg glucose equivalents per endosperm. The tissue residues were suspended in 2 ml of 20 mM sodium phosphate (pH 6.9) and 6 mM NaCl, and boiled for 15 min to gelatinize the starch. Crude boiled homogenates were then used to determine starch according to the method described previously (Hurng and Kao, 1993). Starch level is expressed as mg maltose equivalents per endosperm.
To extract a-amylase, endosperm was homogenized in a chilled (4°C) mortar and pestle with 0.2 M sodium acetate (pH 5.4) containing 3 mM CaCl2. Crude extract was used to determine a-amylase activities by the method developed by Rinderknecht et al. (1967), which uses starch azure as substrate. The change in A595 was used to calculate the a-amylase activity. One unit of enzyme activity is defined as an increase of 1 A595 min-1.
1 Corresponding author.