Bot. Bull. Acad. Sin. (1995) 36: 175-179
Lu et al. Fe- and Mg-deficiency and thylakoid membranes
Influence of Fe- and Mg-deficiency on the thylakoid membranes of a chlorophyll-deficient ch5 mutant of Arabidopsis thaliana
Yih-Kuang Lu1, Yung-Reui Chen1, and Chi-Ming Yang2,3
1 Department of Botany, National Taiwan University, Taipei, Taiwan 106, Republic of China
2 Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 115, Republic of China
(Received January 7, 1995; Accepted May 13, 1995)
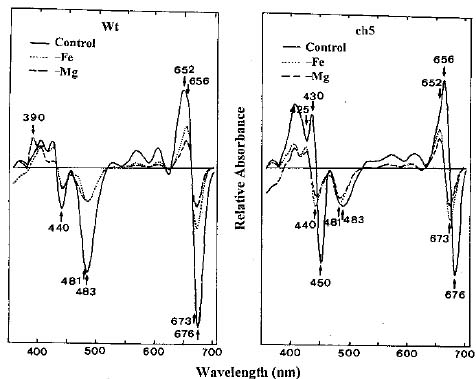
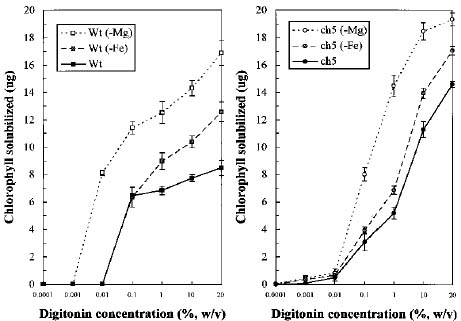
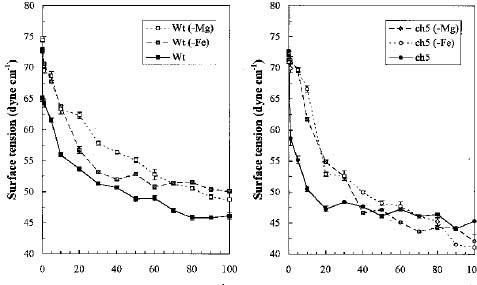
Abstract: We characterized the thylakoid membranes of a chlorophyll-deficient ch5 mutant and a wild type of Arabidopsis thaliana grown in Murashige and Skoog medium with and without Fe or Mg by their composition and surface tension, and by the influence of surfactant on the thylakoid membranes. Deficiency in Fe and Mg decreases the amount of chlorophyll and protein in the ch5 mutant and wild type, but dramatically increases the amount of carotenoid. The pattern of the first-derivative absorption spectra of chloroplast revealed that the composition and organization of thylakoid membranes were very different in the two biotypes. In both biotypes, the surface tension of Fe- or Mg-deficient thylakoid membranes was greater than that in plants grown in normal media, indicating that the thylakoid membranes of Fe- or Mg-deficient plants are more easily accessed by surfactants than those of normally-grown plants. In both biotypes, chlorophyll was more easily solubilized from the thylakoid membranes of Mg-deficient plants than that of Fe-deficient plants, which was more easily solubilized than normally grown plants. We concluded that Fe- or Mg-deficiency results in a looser organization of thylakoid membranes than in the controls, and that Mg-deficiency results in a more disorganized thylakoid membrane than does Fe-deficiency. Results suggest that Fe- and Mg-deficient thylakoids contain smaller amounts of the light-harvesting complexes associated with photosystem II (LHCII) and an abnormal ratio of monogalactosyl diglyceride to digalactosyl diglyceride (MGDG/DGDG).
Keywords: Chlorophyll-deficiency; Fe-deficiency; Light-harvesting complex II; Mg-deficiency; Surface tension; Surfactant; Thylakoid membranes.
Introduction
Iron (Fe) is intimately involved in oxidation-reduction reactions in plants. Magnesium (Mg) is an integral component of the chlorophyll molecule and is a divalent cation cofactor for many enzymatic reactions (Evans and Sorger, 1966). Fe-deficiency causes changes in the structure and composition of thylakoid membranes (Terry and Abadia, 1986; Abadia, 1992), such as a drastic reduction in the amount of thylakoid membrane that has few grana stacks, a general decrease of chlorophyll and carotenoid contents (Stocking, 1975; Pushnik and Miller, 1982; Pushnik et al., 1984; Abadia et al., 1988; Abadia et al., 1989; Guller and Krucka, 1993), a decrease of the ratio of monogalactosyl diglyceride to digalactosyl diglyceride (MGDG/DGDG) (Nishio et al., 1985b), and a depletion of thylakoid membrane proteinsespecially pigment-protein complexes (Nishio et al., 1985a). Plants suffering from Mg-deficiency suffer pronouned changes in the chloroplasts. Photosynthesis, net assimilation, and transpiration rates are decreased under Mg-deficiency (Terry and Ulrich, 1974; Cao and Tibbits, 1992).
The chlorophyll-deficient ch5 mutant of Arabidopsis thaliana was originally induced by ethylmethane-
sulphonate (Koornneef et al., 1983). In a previous study, we examined the composition and biophysical characteristics of thylakoid membranes and concluded that the thylakoid membranes of ch5 mutant were less compact than those of the wild type (Lu et al., 1995a). In a further study, we presented evidence to show that the drastic reduction of light harvesting complex I and II and of MGDG/DGDG ratio affected the grana stacking and swelling (Lu et al., unpublished). This chlorophyll-deficient mutant is able to operate the xanthophyll cycle to respond to the alteration of light intensity, but within a narrower range than does the wild type (Lu et al., 1995b). The aim of our research is to investigate the influence of Fe- and Mg-deficiency on the structure and organization of thylakoid membranes of the ch5 mutant and wild type.
Materials and Methods
Seeds were germinated and grown in Murashige and Skoog (1962) medium for 4 weeks in a growth chamber providing a photoperiod of 12 h L/12 h D, a photoflux of 250_300 mmol m-2 S-1, a relative humidity of 60%, and a temperature of 25°C. Leaves were harvested and the chloroplast and thylakoid membranes isolated as previously described (Markwell, 1986). The concentrations of chlorophyll and carotenoid were determined according to the methods of Arnon (1949) and Jaspers (1965), respectively,
3 Corresponding author.