Bot. Bull. Acad. Sin. (1995) 36: 195-200
Shaw et al. Thermus extracellular a-amylase
Purification and properties of an extracellular a-amylase from Thermus sp.
Jei-Fu Shaw1,2, Fu-Pang Lin1, Su-Chiu Chen2, and Hsing-Chen Chen3
1Institute of Marine Biotechnology, National Taiwan Ocean University, Keelung, Taiwan 202, Republic of China
2Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
3Department of Marine Food Science, National Taiwan Ocean University, Keelung, Taiwan 202, Republic of China
(Received January 4, 1995; Accepted June 5, 1995)
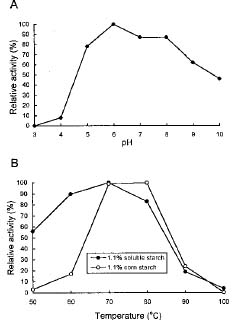
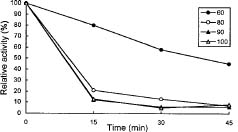
Abstract. An extracellular a-amylase from an extreme thermophile, Thermus sp., was highly purified by affinity absorption on starch granules. SDS-PAGE showed a single band for the purified enzyme, with an apparent molecular weight of 59000. The optimum pH and temperature for the enzyme action on starch was 5.5_6.5 and 70°C, respectively. The enzyme randomly attacked the bonds in the inner region of the starch and produced various maltooligosaccharides. The minimum length of maltooligosaccharide cleaved by this enzyme was maltohexaose. The enzyme activity was strongly inhibited by the addition of Cu2+ and Fe2+ ions. The enzyme belonged to the EDTA-sensitive a-amylase group, but its activity was not stimulated by the presence of Ca2+ ions.
Keywords: Purified; Thermostable extracellular a-amylase; Thermus sp.
Introduction
a-Amylase (EC3.2.1.1, 1,4-a-D-glucan glucano-hydrolase, endoamylase) hydrolyzes starch, glycogen, and related polysaccharides by randomly cleaving internal a-1,4-glucosidic linkages. It is widely distributed in various bacteria, fungi, plants, and animals and has a major role in the utilization of polysaccharides. a-Amylase is an important industrial enzyme. As well as being used as an additive in detergents, it can be used for such things as the removal of starch sizing from textiles, the liquefaction of starch, and the proper formation of dextrin in baking. The thermostability of the a-amylase must be matched to the application. For example, thermostable a-amylases are used for the liquefaction of starch at high temperature and thermolabile a-amylases are used for the saccharification of starch in baking . We have recently developed an enzymatic method that uses thermostable a-amylase in combination with b-amylase and debranching enzymes (isoamylase and/or pullulanase) to produce high-maltose syrup and high protein rice flour simultaneously (Shaw and Sheu, 1992; Shaw, 1994; Shaw et al., 1989).
Thermus is a Gram negative extreme thermophile that produces several thermostable enzymes, such as Taq DNA polymerase (Chien et al., 1976) , pullulanase (Plant et al., 1986; Nakamura et al., 1987), lipase (Phutrakul et al., 1993), and protease (Matsuzawa, 1983), but no report on the production and characterization of a-amylase from Thermus has been published. In this paper, we describe the purification and characterization of a-amylase from a Thermus sp. strain isolated from a hot-spring in northern Taiwan.
Materials and Methods
Screening, Isolation and Identification of Microbial Strains
The sediment samples for screening were obtained from hot-spring sources in Yangmin Shan National Park in northern Taiwan. The organism was enriched on a screening agar plate containing (grams per liter): 2 g (NH4)2 SO4, 0.5 g MgSO47H2O, 0.01 g FeSO47H2O, 0.7 g K2HPO4, 0.3 g KH2PO4, 10 g corn starch, and 17 g agar in distilled water. Incubation at 50_65°C was carried out for 24_48 h, after which the plates were stained with Gram's iodine solution (0.1% I2 and 1% KI) and the colonies with the largest halo-forming zone were isolated for further investigation. Microbiological properties of the isolated strain were determined according to the methods described in Bergey's Manual of Systematic Bacteriology (Sneath et al., 1986).
Amylase Assay
a-Amylase activity was assayed by measuring the reducing sugar released during the reaction, using starch as the substrate, according to the Somogyi-Nelson method. (Nelson, 1944). The reaction mixture contained 50 µl of 1.1% soluble starch (Merck 1252, 0250, MW 6500_8000) in 2 mM imidazole-HCl buffer (pH 7.0) and 250 µl of enzyme solution. The reaction was stopped by adding 100 µl dinitrosalicylic acid solution (100 ml of solution contained 1 g 3,5-dinitrosalicylic acid, 30 g potassium sodium tartarate, and 20 ml 2 N NaOH) after incubation at various temperatures for 1 h. The reaction mixture was then