Bot. Bull. Acad. Sin. (1995) 36: 201-205
Wu and Chen Identification of strain KS3-5
Identification of the Streptomyces strain KS3-5
Rong-Yang Wu1 and Ming-Ho Chen
Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
(Received May 9, 1995; Accepted June 22, 1995)
Abstract. An actinomycete, designated strain KS35, was isolated from a soil sample collected from Kaohsiung, Taiwan, ROC. This organism is capable of producing a series of antibiotics that strongly inhibit the growth of Grampositive and Gramnegative bacteria and yeast-like fungi. The spore morphology and cell wall chemotype suggest that strain KS35 is a streptomycete. Further cultural and physiological characterization and the DNA homology suggest that strain KS3-5 is identical to Streptomyces toxytricini.
Keywords: Streptomyces; Streptomyces toxytricini.
Introduction
The `strike-back' of pathogens has revitalized the search for new drugs (Lemonick, 1994; Jaroff, 1994). Novel antibiotics are required to counter drug-resistant bacteria, fungi, and viruses. Only about 10% of the estimated total number of microbial species are knownthere is an extensive and diverse resource that can be tapped for useful products, such as antibiotics, and processes, such as novel mechanisms of action (Bull et al., 1992). In this respect, natural antibiotics (particularly those from the genus Actinomyces, the most abundant microbial source of anti-microbial compounds; Miyadoh, 1993) are as important as those, such as the b-lactam antibiotics, which are derived from chemical modification of existing antibiotics.
In our screening program for bioactive compounds, an actinomycete (which we designated strain KS35) was isolated from a soil sample collected from southern Taiwan. This actinomycete is capable of producing antibiotics that strongly inhibit the growth of Grampositive and negative bacteria, but appear to be non-toxic to experimental mice and tomato seedlings. These data suggest a use for these KS35 antibiotics in the treatment of animal and plant diseases. We present the identification of strain KS35 through a study of its biological properties.
Materials and Methods
Microorganisms and Culture Conditions
Strain KS3-5 was isolated from a soil sample collected at Tapehu, Kaohsiung, Taiwan. Streptomyces toxytricini ATCC 19813 was purchased from the American Type Culture Collection for comparison. Except where otherwise specified, both strains were cultured on tryptone-yeast extract-glucose (TYG) agar medium containing 10 g glu
cose, 3 g yeast extract, 5 g peptone, 1 g KH2PO4, 1 g K2HPO4, and 20 g agar in 1,000 ml of distilled water, and incubated at 28°C.
Cultural and Morphological Characterization
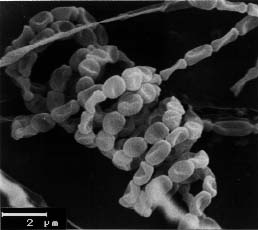
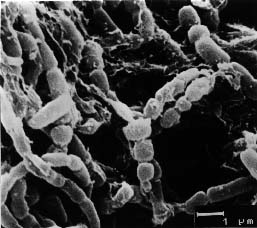
Cultural characteristics of strain KS3-5 were compared on the basis of observations made after 7, 14, and 21 days incubation on Czapek-Dox agar, nutrient agar, Sabouraud agar, and ISP media (Shirling and Gottlieb, 1966). Morphology was examined by light microscopy and scanning electron microscopy (Zeiss DSM model 950).
Physiological Characterization
Utilization of carbohydrates was investigated with a basal carbon nutrient medium (Pridham and Gottlieb, 1948; Waksman, 1967). Methods and media used for physiological tests were as described by Luedemann and Brodsky (1964), Luedemann (1971), Neyra et al. (1977) and Waksman (1967). All cultures were incubated at 28°C for 10 days, except for the gelatin liquefaction (15°C, 21 days). The assay for enzymatic activity was performed according to Hopwood (1967) and Hopwood and Wright (1973). The cultural broth was tested for its antimicrobial activity using the cup or the paper disc diffusion methods (Wu, 1984).
Cell Chemistry
Determination of the cellwall composition, including A2pm (diaminopimelic acid) isomers, sugars, phospholipids, fatty acids and menaquinones was based upon the methods of Becker et al. (1965), Boone and Pine (1968), Kawamoto et al. (1981), Lechevalier and Lechevalier (1970, 1980), and Pine and Boone (1967).
DNA-DNA Homology Study
The DNA-DNA relatedness was determined by the method of Ezaki et al. (1989).
1Corresponding author.