Bot. Bull. Acad. Sin. (1995) 36: 207-214
Yang et al. Coat protein genes of phage Cf
Molecular cloning and expression of the coat protein genes of Cf, a filamentous bacteriophage of Xanthomonas campestris pv. citri
Mei-Kwei Yang1,4, Huei-Mei Huang2, Yen-Chun Yang1, and Wei-Chih Su3
1The Graduate School of Biology, Fu-Jen University, Taipei, Taiwan, Republic of China
2Institute of Life Science, National Defense Medical Center, Taipei, Taiwan, Republic of China
3Graduate Institute of Botany, National Taiwan University, Taipei, Taiwan, Republic of China
(Received April 11, 1995; Accepted August 18, 1995)
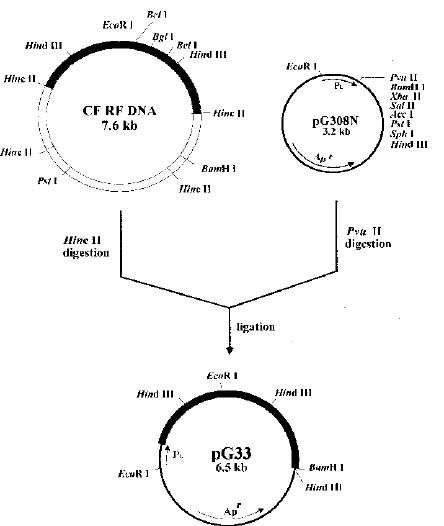
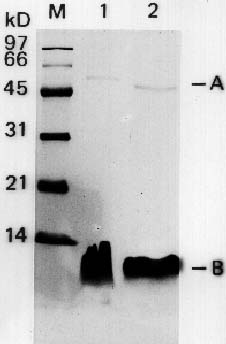
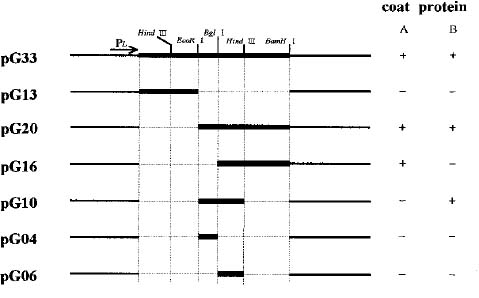
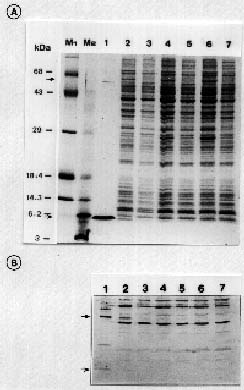
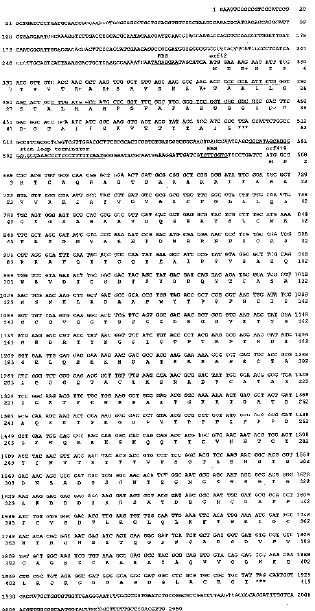
Abstract. Particles of the filamentous bacteriophage Cf contain a major coat protein, the B protein, with a molecular weight of approximately 6,000. In addition, a minor coat protein, the A protein, with a molecular weight of about 50,000, was also identified on sodium dodecyl sulfate-containing polyacrylamide gels. A 3.3 kbp HincII fragment derived from Cf genome was cloned into the expression plasmid pG308N, an E. coli plasmid which carries pL promoter. The recombinant plasmid pG33 and a series of deletion derivatives of pG33 were constructed and transformed into E. coli DG116 for expression of phage Cf genes. The genes coding for A and B proteins of Cf were found on the 2.0 kbp EcoRI-HincII fragment. The complete nucleotide sequences of the 2.0 kbp EcoRI-HincII insert were determined. The deduced amino acid sequence corresponds to a 62-amino acid-residue polypeptide that has a calculated Mr of 6070 was identified as the B protein by SDS/PAGE and immunoblotting. Another open reading frame (ORF419) downstream of the B protein gene (ORF62) was found, and was shown to code for a polypeptide of 419 amino acids with a calculated Mr of 44,676 that exhibits considerable identity to the A protein.
Keywords: Bacteriophage Cf; Coat protein genes; Gene expression; Host specificity; Nucleotide sequences.
Introduction
Cf, Xf, and FLf are filamentous bacteriophages that infect different pathovars of Xanthomonas campestris. All three phages contain a single-stranded circular form DNA surrounded by a long protein coat (Dai et al., 1980; Kuo et al., 1967, 1969; Tseng et al., 1990). Similar to other filamentous phages isolated in Escherichia coli, the infection of these phages results in the releasing of progeny phages into the medium without cell lysis (Hofschneider and Preuss, 1963; Marvin and Hohn, 1969). Although the phages are of comparable genomic size, studies of their host range have revealed a significant difference. Phage Cf, isolated from X. campestris pv. citri, can infect neither X. campestris pv. oryzae nor X. campestris pv. campestris. Another filamentous phage (Xf) isolated from X. campestris pv. oryzae, cannot infect X. campestris pv. citri and X. campestris pv. campestris. We have recently reported, however, that DNAs of Cf and Xf can be transformed into non-host X. campestris strains by electroporation and propagated to accumulate intracellular phage DNA. They then export phage particles at a constant rate (Yang et al., 1991). This demonstrates that adsorption to host cell is responsible for the infectivity of the phage particles. Our interest in the detail of host specificity of phage infection has lead us to examine how Cf adsorbs to X. campestris pv. citri.
The mechanism of adsorption in filamentous phages of E. coli has been inferred from genetic, electron microscopic, and molecular biological studies (Gray et al., 1981; Jacobson, 1972; Rasched and Oberer, 1986; Segawa et al., 1975). It was found that the A protein, specified by gene III of M13, has been implicated in the attachment of the phage to the host receptor (Henry and Pratt, 1969; Pratt et al., 1969), and it may also function in penetration (Goldsmith and Konigsberg, 1977). This adsorption protein comprises a minor fraction of the phage coat and is located only at one end of the phage (Lopez and Webster, 1982; Grant et al., 1981). After infection, the A protein remains with the phage, acting as a pilot protein to guide its DNA into the cell, and converts the single-stranded DNA to the double-stranded replicative form (RF) (Jazwinski et al., 1973; Lin and Bendet, 1976). The A protein also functions as a cut-off agent in the final stage of phage assembly (Crissman and Smith, 1984; Dotto and Zinder, 1983). The adsorption protein of the filamentous phage was found to have both very early and late functions.
Data concerning the structure and function of other coat proteins also exists (Rasched and Oberer, 1986; Simons et al., 1981). B protein encoded by gene VIII is the major protein constituent of the mature phage particles (Henry and Pratt, 1969; Luiten et al., 1983). Within the infected Escherichia coli cell, a large quantities of B protein is synthesized as precursor molecules with amino-terminal signal peptides of 18 to 23 amino acid residues. After insertion of the precursor coat protein into the membrane,
4Corresponding author.