Bot. Bull. Acad. Sin. (1995) 36: 223-228
Lu et al. Amyloplast preparation from sweet potato callus
Preparation of amyloplasts from sweet potato callus culture
Chi-Hui Lu1, Ping-Du Lee1, and Jong-Ching Su1,2,3
1Department of Agricultural Chemistry, National Taiwan University, Taipei, Taiwan, Republic of China
2Institute of Biological Chemistry, Academia Sinica, Taipei, Taiwan, Republic of China
(Received April 7, 1995; Accepted July 27, 1995)
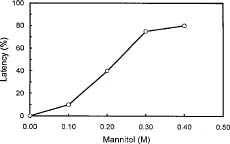
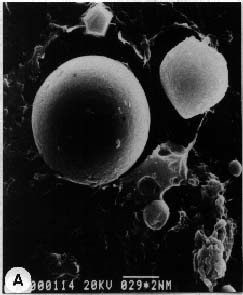
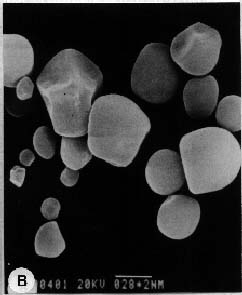
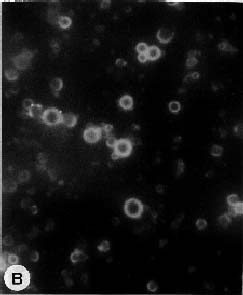
Abstract. Culture conditions for sweet potato callus were manipulated to enhance starch content. Tissue cultures were frozen in liquid nitrogen, pulverized in a ceramic mortar, and ground with a buffer and sea sand. Fractional centrifugation in a discontinuous Percoll gradient enabled separation of amyloplasts in a 40% Percoll layer sandwiched between 20% and 50% layers. A latency test of marker enzymes for amyloplast, and a preliminary feeding experiment with radio-labeled substrates for starch synthesis confirmed that the preparation is suitable for use in the study of starch biosynthesis.
Keywords: Amyloplast; Callus; Ipomoea batatas; Latency test; Starch metabolism; Sweet potato; Tissue culture.
Introduction
Starch synthesis in sink organs occurs in the amyloplasts. The precursor sucrose from the photosynthetic organ must be transformed into appropriate transportable forms to enter the organelle through the amyloplast membrane. Isolated amyloplasts are a desirable research system in which to relate sucrose metabolism in the cytosol to starch synthesis in the amyloplast. To that end, we applied many known methods (Gaynor et al., 1983; McDonald and ap Rees, 1983; Sack et al., 1983; Journet and Dounce, 1985; Macherel et al., 1985; Echeverria et al., 1985; Entwistel and ap Rees, 1988; Mohabir and Hohn, 1988) to tuberous sweet potato root, but were unsuccessful. We noticed that the large size of starch granules in root amyloplasts caused breakdown of the plastid membrane during isolation. To overcome this, we manipulated the culture conditions of sweet potato callus to induce starch synthesis and formation of amyloplasts. We now report our findings.
Materials and Methods
Tissue Culture
Leaves, stems, and young tuberous roots (ranging in size from 1 to 2 cm in diameter) of sweet potato (Ipomoea batatas cv. Tainong 57) were obtained from the University farm just prior to use. The tissue sections for callus initiation were sterilized by immersing in 70% ethanol for 30 sec and in 2.5% NaOCl (containing 1_2 drops of Tween 20 or Triton X-100 wetting agent per 100 ml) for 20 min. Tissue sections (about 5 × 8 × 7 mm) were inoculated on 0.8% agar plates containing 3% sucrose, 2 ppm NAA (1-naphthaleneacetic acid), 0.2 ppm Ki (kinetin), and MS
(Murashige-Skoog) salts and vitamins. Cultures were kept in the dark at 25°C. Callus formation was evident after one week. Subcultures employed MS medium or B5 medium (Gamborg et al., 1982) containing varying concentrations of sucrose, NAA, and Ki. The starch content of tissues was analyzed at three-week intervals. Growth in a culture medium was estimated by the change in dry weight of tissues.
Enzyme Extraction
The callus tissue was mixed with sea sand, frozen in liquid nitrogen, and ground to a powder with a mortar and pestle. One to one-and-a-half times its weight of a 20 mM HEPES buffer (pH 7.8) containing 2 mM KCl, 2 mM CaCl2, 1 mM EDTA, and 1% polyvinylpolypyrrolidone (PVPP) was added and the mixture was homogenized at 2°C for one min. After centrifugation at 10,000 g the supernatant liquid was used for assays of proteins (Bradford, 1976), SP (starch phosphorylase) (DeFekete and Vieweg, 1974), ADPGPPase(ADPG pyrophosphorylase) (Nakamura et al., 1989), PPase (inorganic pyrophosphatase) (Gross and ap Rees, 1986), SS (sucrose synthase) (Su et al., 1977), and amylase (Street, 1974).
Amyloplast Isolation
About 50 g of callus was rapidly frozen in liquid nitrogen and pulverized with a mortar and pestle. The powder was slowly added (with gentle agitation, avoiding freezing) to 500 ml of 50 mM HEPES (pH 7.8) buffer containing 5 mM MgCl2, 2 mM EDTA, 0.1% bovine serum albumin, 5 mM DTT (dithiothreitol), 1 mM CaCl2, and 0.3 M mannitol (buffer A). Subsequent operations were performed at 4°C. The suspension was filtered in succession through 300 µm and 64 µm nylon mesh. The filtrate was centrifuged at 190 g for 6 min and the supernatant liquid was discarded. The precipitate was suspended in buffer A, then Percoll was added to 30%. Three milliliters of
3Corresponding author.