Bot. Bull. Acad. Sin. (1995) 36: 247-251
Hsiao and Lin Chemotaxonomy of Clerodendrum
A chemotaxonomic study of essential oils from the leaves of genus Clerodendrum (Verbenaceae) native to Taiwan
J. Y. Hsiao1 and M. L. Lin
Department of Botany, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received January 11, 1995; Accepted July 11, 1995)
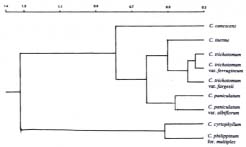
Abstract. Fifty-four mature-leaf samples of nine Clerodendrum taxa native to Taiwan were collected from various locations on the island. Each sample was steam-distilled, and the resulting essential oil was analyzed by gas chromatography. The relationship among taxa was analyzed by cluster analysis of the gas chromatogram data (463 peaks). We assumed that peaks with the same retention time on different chromatograms were the same compound. The cluster analysis indicated a congruence between morphological and chemical relationships at the intraspecific level. All intraspecific taxa were linked as distinct clusters. At the interspecific level, there was congruence and discrepancy between morphological and chemical relationships. The results of the present study support the taxonomic treatment in which the white-flowered variant of C. paniculatum is recognized as a variety of the species. The results also support the existence of C. trichotomum var. fargesii and C. trichotomum var. ferrugineum as intraspecific taxa of C. trichotomum.
Keywords: Chemotaxonomy; Clerodendrum; Essential oils; Taxonomy; Verbenaceae.
Introduction
The genus Clerodendrum (Verbenaceae) is widely distributed in the tropics and subtropics, with a few species extending into the temperate regions. The species number has been estimated to be five hundred and sixty (Moldenke, 1971) and five hundred and eighty (Munir, 1989). The genus was first described by Linnaeus in 1753, based on the type species Clerodendrum infortunatum from India. In 1763, Adanson changed the Latinized form, Clerodendrum, into the Greek form, Clerodendron. In 1942, Moldenke re-adopted Linnaeus' original Latinized form, and the practice has been followed by most taxonomists. The genus comprises small trees, shrubs (sometimes climbing), and (rarely) herbs. Morphologically it is characterized by opposite and exstipulate leaves, tetragonal or terete stems, terminal or axillary cymose inflorescence or panicles, hypogynous bisexual flowers, persistent calyx, cylindrical corolla-tube with spreading 5-lobes at the top, epipetalous and exserted stamens, long-exerted style and short-bifided stigma, imperfectly 4-celled ovary, exalbuminous seeds, and endocarp separating into 2 to 4 stony pyrenes.
The species of Clerodendrum native in Taiwan are distributed from the coasts to low-to-middle altitudes in the mountains. The treatments by taxonomists are diverse (e.g. Hayata, 1912; Kanehira, 1917; Li, 1963; Moldenke, 1971; Hsiao, 1978; Liu and Liao, 1981; Hue and Chen, 1982; Liu et al., 1988). The indisputable taxa includes
C. canescens Wall., C. cyrtophyllum Turcx., C. inerme (L.) Gaertner, and C. philippinum Schauer forma multiplex (Sweet) Moldenke. Among the more controversial taxa, several authors (Moldenke, 1971; Hsiao, 1978; Liu et al., 1988) treated C. trichotomum Thunb. as comprising three varieties, while others (Li, 1963; Liu and Liao, 1981) recognized the existence of only two varieties, with the exclusion of C. trichotomum var. ferrugineum. Hue and Chen (1982) recognized the species without any intraspecific division. Another controversial taxon is the white-flowered variant of C. paniculatum L., which was published as a new variety by Hemsley in 1895 and named C. paniculatum var. albiflorum Hemsley. Hsieh (1973), however, lowered its rank to a form and named it C. paniculatum L. forma albiflorum (Hemsl.) Hsieh. The existence of C. intermedium Cham. in Taiwan is questionable even though it was treated as a native species by Hsiao (1978) and Liu (1981). In the present study, a plant close to the description of C. intermedium, and having leaves without lobes, was collected from Kenting Botanic Garden (Lin 1050). This solitary specimen is thought to be a cultivated plant, and this species is not included in the present study.
The information gathered from essential-oil studies has proved to be of value in the taxonomic and evolutionary investigations of plants (Von Rudloff, 1975; Southwell and Stiff, 1990; Brophy and Clarkson, 1992; Brothy et al., 1994). The purpose of the present study is to re-evaluate the relationships among native taxa of Clerodendrum in Taiwan, based on their essential-oil compositions. It is expected that these data will shed light on the natural relationships of the taxa.
1Corresponding author.