Bot. Bull. Acad. Sin. (1996) 37: 1-7
Lin and Yao Mungbean protease activities
Mungbean (Vigna radiata L. Wilczek) contains some high proteolytic activities already before germination
Yaw-Huei Lin1 and Wen-Hsiang Yao
Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
(Received January 13, 1995; Accepted September 8, 1995)
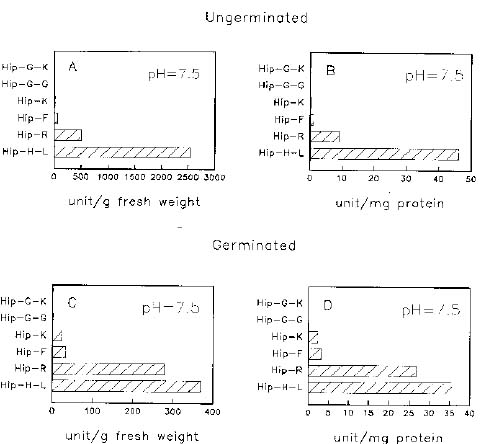
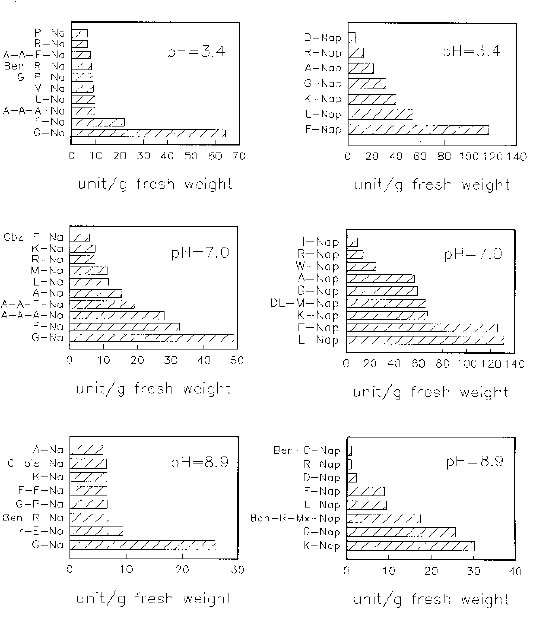
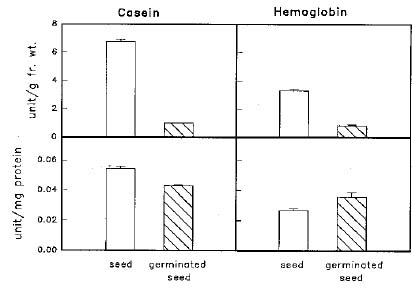
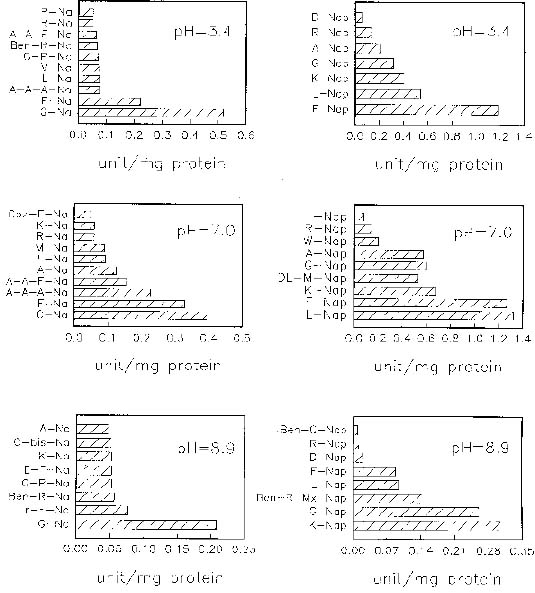
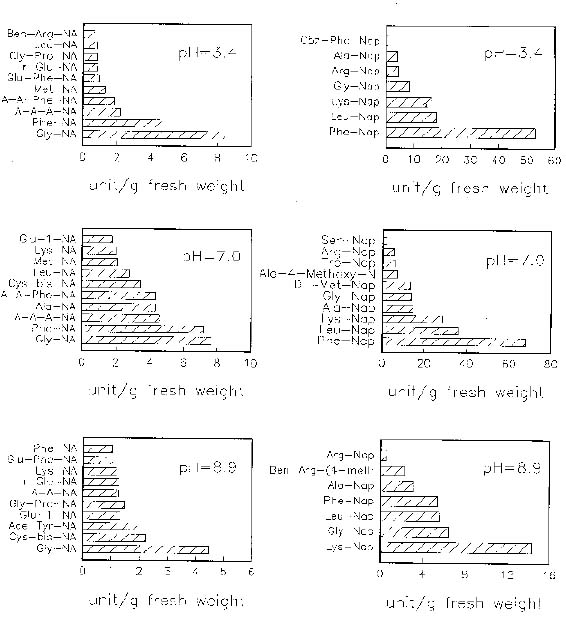
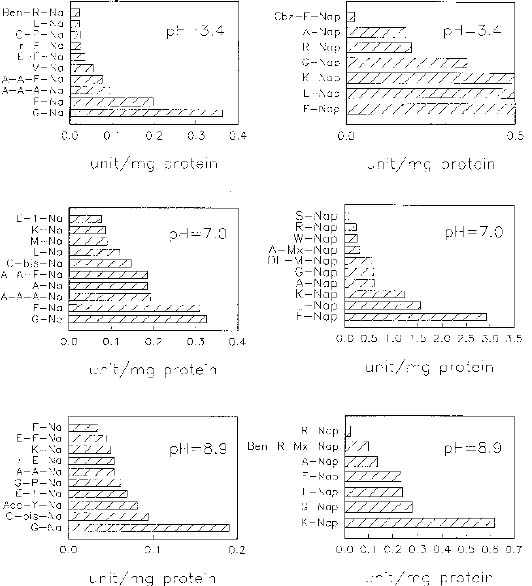
Abstract. When casein was used as a substrate, mungbean seeds had higher proteinase activity levels based on g fresh weight or on mg protein before germination than after germination. While hemoglobin was used, the same was observed as based on g fresh weight but the opposite was observed as based on mg protein. Carboxypeptidase activities toward Hip-His-Leu or Hip-Arg at pH 7.5 based on g fresh weight were also higher before germination than after germination, but activity toward Hip-Arg based on mg protein before germination was lower than after germination. Before germination, when 4-nitroanilides were used, Gly-Na and Phe-Na were hydrolyzed most rapidly at pH 3.4. Gly-Na, Phe-Na, Ala-Ala-Ala-Na, and Ala-Ala-Phe-Na were hydrolyzed most rapidly at pH 7.0. Gly-Na and g-Glu-Na were hydrolyzed most rapidly at pH 8.9. When b-naphthylamine derivatives were used, Phe-Nap, Leu-Nap, and Lys-Nap were hydrolyzed most rapidly at pH 3.4 while Leu-Nap and Phe-Nap were hydrolyzed most rapidly at pH 7.0. Lys-Nap, Gly-Nap, and Ben-Arg-Mx-Nap were hydrolyzed most rapidly at pH 8.9. After germination, when 4-nitroanilides were used, Gly-Na, Phe-Na, Ala-Ala-Ala-Na, and Ala-Ala-Phe-Na were hydrolyzed most rapidly at pH 3.4. Gly-Na and Phe-Na were hydrolyzed most rapidly at pH 7.0. Gly-Na was hydrolyzed most rapidly at pH 8.9. When b-naphthylamine derivatives were used, Phe-Nap, Leu-Nap, and Lys-Nap were hydrolyzed most rapidly at pH 3.4 and 7.0. Lys-Nap and Gly-Nap were leading substrates at pH 8.9. Mungbean enzymes hydrolyze derivatives of 4-nitroaniline and b-naphthylamine of amino acids and peptides differently at various pH values.
Keywords: Protease; Protein substrates; Synthetic substrates; Germination; pH.
Abbreviations: Ace, acetyl; Ben, benzoyl; Cbz, benzylcarbonyl; Cp, N-3-(carboxy propionyl)-; Hip, hippuryl; Mx, methoxy; Na, nitroanilide; Nap, naphthylamide.
Introduction
There are many reports of plant protease activities in the literature. Most reports are on legumes (Citharel and Garreau, 1987; Couton et al., 1991; Collier and Murray, 1977; Crump and Murray, 1979; Dunaevsky and Belozersky, 1989; Elleman, 1974; Mikkonen, 1986; Mikkonen, 1992; Wong, 1991; Wynn and Murray, 1985; Yamaoka et al., 1990; Yamaoka et al., 1994; Yu and Greenwood, 1994); some are on crops (Horiguchi and Kitagishi, 1976; Kolehmainen and Mikola, 1971; Vodkin and Scandalios, 1980; Waters and Dalling, 1983); some on vegetables (Kitamura and Maruyama, 1985; Lin and Chan, 1990); and a few reports are on others (Ninomiya et al., 1981; Pallavieni et al., 1981).
Protease activities in a variety of plants are reported in two books edited by Dalling (1986). Aspartic proteinase and some amino peptidase activities are present in ungerminated seeds. Some of these enzymes have been purified and cloned (Kolehmainen and Mikola, 1971; Ninomiya et al., 1981, 1983; Runeberg-Roos et al., 1991; Salmia and Mikola, 1975; Sarkkinen et al., 1992). Cysteine proteinase (Shutov and Vaintraub, 1987) and
carboxypeptidases (Dunaevsky et al., 1987) are expressed in germinating and post-germinating seeds. Amino peptidases are expressed in sprouts of sweet potato (Lin and Chan, 1992). There are two major storage proteins in soybean (Glycine max [L.] Merrill) seed, glycinin and b-conglycinin. During germination and early growth, these storage proteins are degraded by proteolysis. The predominant pattern is one of limited proteolytic cleavage by proteases specific to the reserve protein, followed by more rapid proteolysis by less specific proteases (Muntz et al., 1985; Shutov et al., 1982). Several soybean proteases have been described, including six proteolytic enzymes from ungerminated seeds separated by anion-exchange chromatography (Wei et al., 1966), two carboxypeptidases from germinating soybean (Kubota et al., 1976), two endopeptidases, one exhibiting an acidic pH optimum and the other a basic pH optimum (Bond and Bowles, 1983), and a trypsin-like protease (Nishikata, 1984).
Many reports also deal with mungbean, especially with cotyledons of mungbean. Results have suggested that the appearance of a sulfhydryl-type endopeptidase activity is a necessary prerequisite for the rapid metabolism of the reserve proteins which accompanies the germination of mungbean (Chrispeels and Boulter, 1975). Results have also suggested that new enzymatic activities not present
1Corresponding author.