Bot. Bull. Acad. Sin. (1996) 37: 17-23
Chan et al. cryIA(a) gene expression in transgenic potato
Expression of Bacillus thuringiensis (B.t.) insecticidal crystal protein gene in transgenic potato
Ming-Tsair Chan1,4, Liang-Jwu Chen2, and Hsin-Hsiung Chang3
1Institute of Molecular Biology, Academia Sinica, Nankang, Taipei, Taiwan 11529, Republic of China
2Institute of Molecular Biology, National Chung Hsing University, Taichung, Taiwan, Republic of China
3Department of Agronomy, National Taiwan University, Taipei, Taiwan 10764, Republic of China
(Received May 04, 1995; Accepted August 29, 1995)
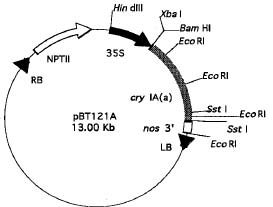
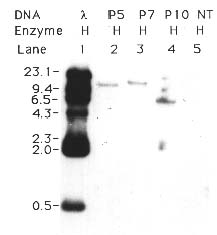
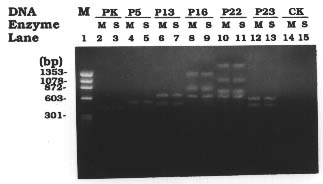
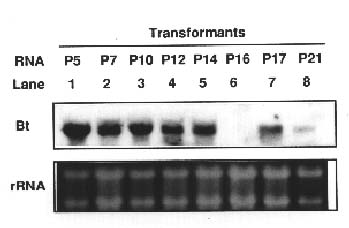
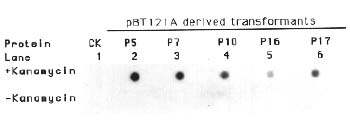
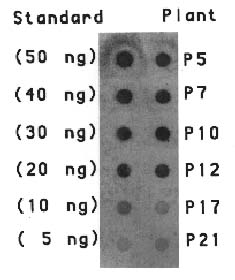
Abstract. The crystal proteins, d-endotoxins, of Bacillus thuringiensis are specifically lethal to Lepidopteran insects. A truncated B.t. toxin gene, cryIA(a), encoding an insecticidal crystal protein (ICP) directed by the cauliflower mosaic virus 35S promoter was transferred to potato plants by an Agrobacterium-mediated transformation system. The integration of the cryIA(a) gene into potato genome was determined by Southern blot analysis and polymerase chain reaction (PCR). The copy number of the integrated gene was estimated by inverse polymerase chain reaction (IPCR). The cryIA(a) RNA transcripts in transgenic potato plants were demonstrated by Northern blot analysis. Seven out of thirty transgenic plants expressed the cryIA(a) gene. Those transgenic plants containing multiple transgene copies did not express cryIA(a) gene. Nevertheless, transgenic potato plants grown in the greenhouse contained 7_52 ng ICP per gram fresh leaf.
Keywords: Agrobacterium tumefaciens; Bacillus thuringiensis; cryIA(a) gene expression; Insecticidal crystal protein; Transgenic potato.
Abbreviations: ICP, insecticidal crystal protein; IPCR, inverse polymerase chain reaction; PCR, polymerase chain reaction; PSC, potato suspension cultures; RB, T-DNA right border; LB, T-DNA left border; NPTII, neomycin phosphotransferase II.
Introduction
The potato (Solanum tuberosum L.) is one of the major crops in agricultural production. Current efforts to develop insect-resistant crops through biotechnology are based primarily on transforming plants with a single gene encoding insecticidal enzyme or toxin. The most widely used genes in this approach are the d-endotoxin gene of Bacillus thuringiensis, a sporeforming, gram-positive bacterium. The insecticidal crystal protein (ICP) from the B. thuringiensis var. kustaki is a specific toxin for lepidopteran insects yet exhibits no toxicity toward humans, other vertebrates, or beneficial insects (Delannay et al., 1989). Formulated bacterial products have been used as insecticides for a long time. However, practical usages of such microbial products are limited because of their relatively high cost and poor persistence under field conditions, resulting in a need for multiple applications (Sneh et al., 1983).
Lepidopteran-active ICPs are protoxins of MW. 130_160 kDa. These protoxins emerge when exposed to an alkaline medium (pH 9_12), such as that found in the insect midgut. These protoxins are proteolytically cleaved into smaller, active forms (MW 60_70 kDa) derived from
the N-terminal half of the protein (Hofte et al., 1989). Although the mode of toxin action is largely unknown, it is assumed to bind specific proteins on the membrane of the insect gut (Hofmann et al., 1988).
The B.t. toxin cryIA gene of B. thuringiensis has been engineered and transferred into several plant species to yield resistance against certain lepidopteran insects. The truncated genes, which produce insecticidally active protein, have been expressed in potato (Adang et al., 1993; Perlak et al., 1993), tomato (Delannay et al., 1989), tobacco (Barton et al., 1987; Fischhoff et al., 1987; Vaeck et al., 1987), cotton (Perlak et al., 1990), corn (Koziel et al., 1993), and rice (Fujimoto et al., 1993). The use of a native d-endotoxin coding region, which has a high A-T content, appears to lead to an abnormally low expression in plants. Modifications of the coding region sequence to increase the G-C content of the native gene resulted in a dramatic increase in the expression of the insecticidal protein (Perlak et al., 1991).
As a first step toward the development of an insect resistant potato, attempts were made to transfer the truncated cryIA(a) gene directed by the cauliflower mosaic virus 35S promoter into potato plants through Agrobacterium-mediated transformation. These transgenic potato plants could provide alternatives to hazardous synthetic chemical insecticides for controlling lepidopteran pests.
4Corresponding author.