Bot. Bull. Acad. Sin. (1996) 37: 35-40
Chang and Yang Embryo culture of Taxus mairei
Enhancement of plant formation from embryo cultures of
Taxus mairei using suitable culture medium and PVP
S.H. Chang1 and J.C. Yang
Taiwan Forestry Research Institute, 53 Nan-Hai Road, Taipei, Taiwan, Republic of China
(Received May 23, 1995; Accepted September 29, 1995)
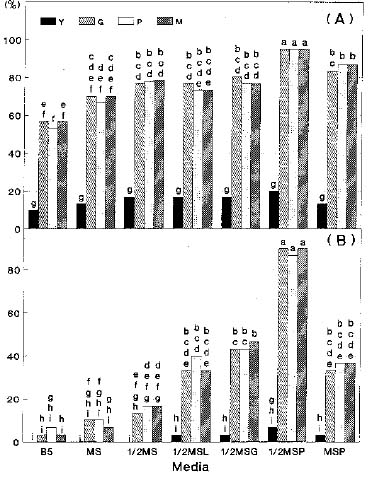
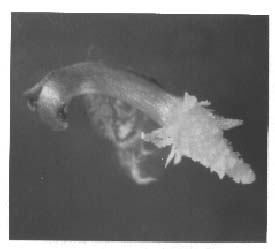
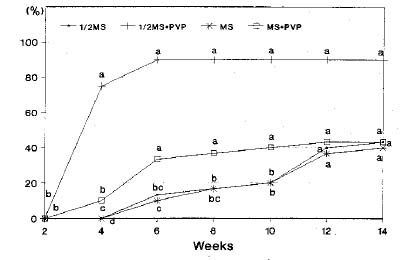
Abstract. Seedlings of Taxus mairei, of which seeds have proved difficult to germinate, were obtained by embryo cultures. Seed development in four stages was classified according to the visible characteristics of the fruits; viz, young fruits without aril, green fruits with undeveloped green arils, pink fruits with enlarging pink arils and mature fruits with fully developed red arils. The composition of the medium was the critical determinant. A 1/2MS medium with PVP supported the best embryonic growth and seedling development not only for T. mairei but also for T. baccata, T. canadensis, T. cuspidata and T. media. Only 15% of young embryos of T. mairei, in this medium, developed into plants. However, nearly ninety percent of embryos at green, pink, and mature stages developed into plants. A 90% survival rate of in vitro plantlets grown in the greenhouse was obtained.
Keywords: Immature embryo culture; Mature embryo culture; PVP; Taxus mairei.
Abbreviations: GA3, gibberellin acid; PVP, polyvinylpyrrolidone-40.
Introduction
Taxol, an effective anticancer drug for advanced ovarian and other cancers (NCI, 1990), has been found in the bark, root, and other parts of Taxus brevifolia (Wani et al., 1971). Other Taxus species have been extensively examined as additional potential sources due to the increasing demand for taxol (Fett-Netto et al., 1992). Taxus mairei, sparsely distributed in northern and central parts of Taiwan at altitudes of 1,000_3,000 m (Liu, 1980), is a slowly growing tree that reaches heights of up to 17 m and possesses good wood quality for furniture and sculpture. The species faces the threat of extinction because of strong demand for both wood and taxol production and difficulty in propagation. Most of the fruits are eaten by birds and are difficult to collect. Furthermore germination of mature seeds required a series of treatments. Alternative temperatures and cold stratification for 8.5 months produced a 50% germination rate (Chien et al., 1994). Vegetative propagation then is an alternative (Sheu, 1985); however, strong topophysis of lateral shoots limited its application for the production of planting stocks.
Embryo culture is a useful tool to overcome seed dormancy and to abbreviate the breeding cycle (Ho, 1987). Embryonic growth was first observed from cultures of immature and mature embryos of T. baccata (Le Page-Degivry, 1968; Le Page-Degivry and Garello, 1973). Immature embryo cultures, thought to contain less inhibitory compounds than mature embryos, were carried out in T. baccata, T. brevifolia, T. cuspidata and T. media. The fraction of seedling formation from immature
embryos, however, was 15 to 40% (Flores and Sgrignoli, 1991). Mature embryo cultures were difficult to develop into seedlings (Flores and Sgrignoli, 1991; Hu et al., 1992). Many treatments to increase germination viz: GA3 added into the culture medium, a 14 h photoperiod during incubation, and an initial four weeks of culture in darkness were reported to improve embryo germination and growth into seedlings for T. brevifolia, T. baccata and T. cuspidata (Hu et al., 1992; Flores et al., 1993; Chee, 1994). The objective of our study was to find an optimal medium through which embryo dormancy could be bypassed and mass-production of T. mairei seedlings could be facilitated for conservation planting and taxol production.
Materials and Methods
Seed Material
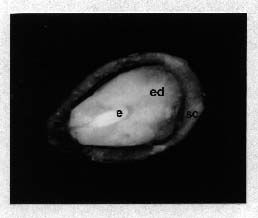
Taxus mairei fruits were collected from Nan-Chuang, in the north-western part of Taiwan, ROC. Mature fruits were gathered in December 1992 and 1993. Immature fruits were collected from late September to December 1993. Seeds were sorted into the following groups according to the visible developmental characteristics of the fruits and aril: Y, young seeds from early gathered fruits without aril development; G, seeds from green fruits with undeveloped green aril; P, seeds from pink fruits with enlarging pink aril and M, mature seeds from red fruits with fully developed red aril. Fruits were rubbed over screens, washed thoroughly, and the light seeds were rinsed off. All seeds were mixed with wet sphagnum, placed in closed polyethylene bags, and stored in a refrigerator at 4°C. Fresh seeds were cultured within two weeks after collection. For comparison of storage effect, 6-month-old
1Corresponding author.