Bot. Bull. Acad. Sin. (1996) 37: 41-49
Wu et al. 5' Flanking region of glutelin genes
Analysis of 5' region of glutelin genes from wild rice species
Hsin-Kan Wu1, Tein-chin Chen, and Mei-Chu Chung
Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan, Republic of China
(Received June 7, 1995; Accepted September 13, 1995)
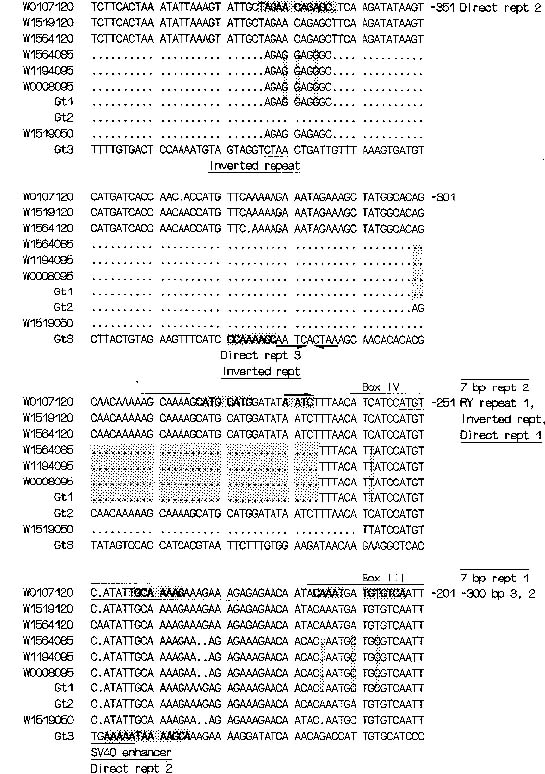
Abstract. The structure of the 5' flanking region of glutelin genes amplified from the various wild rice species was analyzed by cloning and sequencing. The results showed that beyond the essential boxes (legumin, CAAT, AACA and TATA), the 5' region of rice glutelin genes have numerous putative enhancers (long-direct and short-direct repeats) and putative regulatory segments (RY repeats, -300 bp elements, nuclear protein binding sites) though portions of a few elements have been deleted in some wild species. The possible roles of most of the putative elements in glutelin gene expression remain to be determined. The sequence length and structure of glutelin 5' regions vary among rice species. On the basis of the length, the degree of homolgy, and the corresponding base substitutions and deletions in the 5' regions of glutelin genes, the authors suggest that glutelin genes, in the subfamily Glua can be classified into three kinds of members, each with its 5' region of 0.5 kb, 0.9 kb, or 1.2 kb. A member gene in the subfamily may reside at one or more loci in a rice genome. The same member gene that appears among rice species with minor deletion, addition, or substitution may be designated as alleles of that gene.
Keywords: Glutelin gene; 5' Region sturcture; Subfamily GluA; Wild rice.
Introduction
A rice glutelin cDNA was first isolated from cultivated rice by Takaiwa et al. (1986). More glutelin cDNAs were isolated and classified into two types, I and II, which can be distinguished respectively by stop coden TAA or TAG and by two and one polyadenylation signals (Takaiwa et al., 1987a). Since these early reports, three genomic clones (Gt1, Gt2 and Gt3) for rice glutelin have been isolated and studied in Okita's laboratory. Comparison of DNA sequences from relevant regions of these clones showed that two of them, Gt1 and Gt2, are closely related. Gt3 shows little or no homology to Gt1 and Gt2. All three clones had 5' flanking regions of less than 0.9 kb (Okita et al., 1989). Two new glutelin genes, Glua-3 and Glua-4 were later added to subfamily A (Takaiwa and Oono, 1991) that already contains Glua-1 (Type I) and Glua-2 (Type II). Takaiwa et al. (1991) proposed a new subfamily of glutelin gene, subfamily B, in which three member genes have been sequenced. Furthermore, direct repeat, enhancer core, and legumin box (Takaiwa et al., 1987b), -300 bp element, RY repeats and inverted repeats (Okita et al., 1989) and nuclear protein binding sites (Kim and Wu, 1990; Takaiwa and Oono, 1990) have been reported to be related to glutelin gene expression in cultivated rice. This paper reports the structure of 5' regions of glutelin genes with special emphasis on that of glutelin subfamily A genes from wild rice species.
Materials and Methods
The species, both cultivated and wild, used in this experiment are shown in Figure 1.
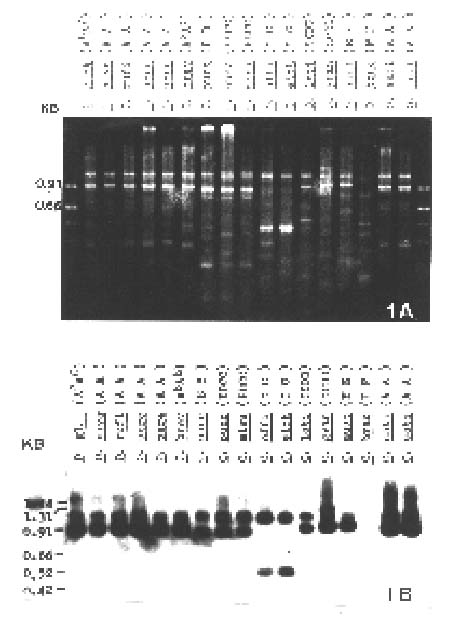
Figure 1. Southern blot analysis of the amplified rice glutelin 5' region. 1A, Gel electrophoresis of the PCR amplified 5' regions of glutelin genes from various rice species each indicated above the line. Each has two to three major bands; 1B, The blotted DNA was probed with the 5' region of a glutelin gene, isolated from rice cultivar Tainung 67 (genome AA). It shows that all wild rice species except O. brachyantha (genome FF) have two major bands positively reacted.
1Corresponding author.