Bot. Bull. Acad. Sin. (1996) 37: 93-98
Weng et al. Plasmid from Xanthomonas
Characterization of the 1.8-kb plasmid pXV64 from Xanthomonas campestris pv. vesicatoria
Shu-Fen Weng1,4, Nien-Tsung Lin2, Yu-Fen Fan1, Juey-Wen Lin3, and Yi-Hsiung Tseng1
Institute of Molecular Biology1, Department of Botany2 and Agricultural Biotechnology Laboratories3, National Chung Hsing University, Taichung 402, Taiwan, Republic of China
(Received October 7, 1995; Accepted January 31, 1996)
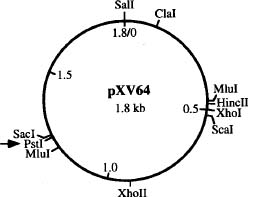
Abstract. A simple rapid plasmid-screening method was developed and then used on 58 strains of Xanthomonas campestris pv. vesicatoria. A very small (1.8-kb) cryptic plasmid, designated pXV64, that had a copy number of about 200 to 350 was detected. A physical map of pXV64 was established and a derivative, designated p64PK, was constructed by cloning a kanamycin resistance cartridge into the unique PstI site of pXV64. p64PK had the ability to transform different strains of Xanthomonas, could be maintained stably, and was compatible with the broad-host-range IncP plasmid pLAFR1. The results of this study suggest that pXV64 has the potential to be developed into vectors for gene cloning in Xanthomonas.
Keywords: Plasmid; Rapid plasmid-screening method; Vector; Xanthomonas campestris.
Introduction
Xanthomonas campestris, consisting of more than 123 pathovars, is the major species of the genus Xanthomonas, a genus that includes many gram-negative plant-pathogenic bacteria (Vauterin et al., 1990). Xanthomonas campestris pv. vesicatoria (Xv) is the pathogen for foliage and fruit spot disease in peppers and tomatoes (Minsavage et al., 1990). Xanthomonas campestris pv. campestris (Xc) is the causal agent of black rot in crucifers (William, 1980).
While plasmids from several pathovars of Xanthomonas have been detected, detailed studies on these plasmids are limited (Lin et al., 1979; Tetaz and Luke, 1983; Stall et al., 1984; Civerolo, 1985; Stall et al., 1986; Lazo and Gabriel, 1987; Chen and Tseng, 1988; Xu and Gonzalez, 1991). Efforts to develop plasmids indigenous to Xanthomonas into cloning vectors have yet to be reported. In this study, we developed a rapid method for screening plasmids in 58 strains of X. campestris pv. vesicatoria, which resulted in detection of a miniplasmid pXV64 in strain Xv64. As this plasmid is very small, stably maintained at a high copy number, and is compatible with IncP plasmid, it has good potential to be developed into a shuttle vector. Preliminary tests carried out in this study have demonstrated this possibility.
Materials and Methods
Bacterial Strains, Plasmids and Culture Conditions
Fifty-eight strains of X. campestris pv. vesicatoria, X. campestris pv. citri, X. campestris pv. phaseoli, X.
campestris pv. begoniae, X. oryzae pv. oryzae, and Rhizobium japonica RJ119 were obtained from S. T. Hsu, National Chung Hsing University. Xanthomonas campestris pv. campestris 17 (Yang and Tseng, 1988) and X. campestris pv. campestris P20H (Yang et al., 1988) came from our laboratory collection. Other strains used were Agrobacterium tumefaciens LBA4404 (Clontech Laboratories, Inc.), Azotobacter vinelandii ATCC478 and Pseudomonas putida IF03738 (ATCC8209). Luria broth or L agar (Miller, 1972) was used to grow E. coli HB101 (Boyer and Roulland-Dussoix, 1969) at 37°C or all other bacteria at 28°C. Antibiotics used were: ampicillin, 50 µg/ml; chloramphenicol, 34 µg/ml; kanamycin (Km), 50 µg/ml; and tetracycline (Tc), 15 µg/ml.
The plasmids used were pXV64, pXV2 (14.6 kb, our laboratory collection), pRK415 (10.5 kb, Tcr; Keen et al., 1988), pK701 (15.8 kb, pRK415 carrying a 5.3-kb fragment from Xc17 chromosome), pBR325 (6.0 kb; Bolivar, 1978), pACYC184 (4.2 kb, Chang and Cohen, 1978), and pUC18 (2.7 kb; Yanisch-Perron et al., 1985).
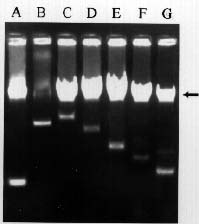
Rapid Screening for Plasmids
A small number of cells were picked by touching a colony with a toothpick. They were then inoculated into a microfuge tube containing 300 µl of L broth. After overnight growth, the cells were pelleted in a microfuge (10,000 rpm, 2 min). The cells were then suspended by vortexing in 20 µl of gel-loading mix (0.25% bromophenol blue and 30% glycerol). Then 40 µl each of chloroform and phenol (saturated with 1.0 M Tris-HCl, pH 8.0) was added. The mixture was vortexed at full speed for 1 min followed by centrifugation for 10 min at 12,000 rpm. Then 10 µl of the aqueous fraction was subjected to electrophoresis on 0.7% agarose minigel (5.2 × 6.0 cm) with TAE buffer (40 mM Tris-acetate, pH 8.0 containing 2 mM Na2-EDTA) at 100 volts for 30 min. The gel was stained
4Corresponding author. Fax: 886-4-2874879.