Bot. Bull. Acad. Sin. (1996) 37: 99-105
Chen et al. Nucleotide sequence of the nifHDK operon
Nucleotide sequence of the nifHDK operon in the aerobic
nitrogen-fixing unicellular Synechococcus RF-1
Horng-Ming Chen1,2, Tan-Chi Huang2,3, and Chiu-Yuan Chien1
1Department of Biology, National Taiwan Normal University, Taipei, Taiwan 117, Republic of China
2Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 115, Republic of China
(Received February 5, 1996; Accepted March 19, 1996)
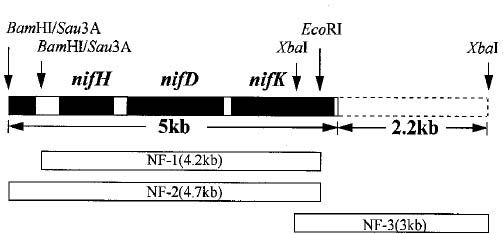
Abstract. A 5 kb nucleotide sequence which included nifHDK operon was determined. The amino acid sequences deduced from the nifH, nifD, and nifK of Synechococcus RF-1 and from Anabaena 7120 were compared. The start site of transcription was located by primer extension analysis. A region of 405 bp just upstream from the 5' end of nifH showed considerable sequence similarity to the nifU sequences of Anabaena 7120.
Keywords: Cyanobacteria; nifHDK operon; Nitrogen fixation; Nucleotide sequence; Synechococcus RF-1.
Introduction
The nitrogenase which catalyzes the reduction of atmospheric nitrogen to ammonia is a complex consisting of three different kinds of polypeptide encoded by nifH, nifD, and nifK, respectively. The molecular structure of nifHDK operon has been analyzed in eubacteria, photobacteria, and cyanobacteria. The nucleotide sequence for the coding region of nif genes among different microorganisms is highly conserved. However, the arrangement of nifH, nifD, and nifK is either in a contiguous or noncontiguous array. The nifH, nifD, and nifK in the eubacteria system such as in Klebsiella pneumoniae (Roberts and Brill, 1981), or Rhizobium meliloti (Corbin et al., 1982), and in the non-oxygenic photosynthetic bacteria, such as in Rhodopseudomonas capsulata (currently, Rhodobacter capsulatus), are linked in a single nifHDK operon (Avtges et al., 1983). In the cyanobacteria, two types of arrangement are found. Those in the nonheterocystous anaerobic nitrogen-fixing cyanobacteria such as in the Synechococcus 7335, Synechococcus 7425, or Pseudoanabaena 7409 are linked in a contiguous array (Kallas et al., 1985). However, those in heterocystous cyanobacteria such as in Anabaena 7120, nifH is linked to nifD, whereas nifK is separated from nifD by about 11 kb in the vegetative cells (Mazur et al., 1980). A rearrangement of the nif genes takes place during the initiation of heterocyst development (Golden et al., 1985).
Among the unicellular cyanobacteria, a few isolates are able to fix nitrogen aerobically. They can be morphologically grouped into sheathed and sheathless forms (Waterbury and Rippka, 1989). These aerobic nitrogen-fixing cyanobacteria fix nitrogen almost exclusively during the dark periods when grown in a diurnal 12 h/12 h light-dark regimen. The properties of the nitrogenase activity in Synechococcus RF-1, one of the sheathless aerobic nitro
gen-fixing unicellular cyanobacteria have been well characterized. When the culture was exposed to a diurnal 12 h light/12 h dark (L/D) regimen, the nitrogenase activity exhibited a rhythmic pattern with the nitrogenase activity peak occurring in the dark phase. This rhythmic pattern persisted for several days after the culture was transferred to continuous light (L/L) (Grobbelaar et al., 1986). The nitrogenase activity rhythm was also found to be temperature compensated when the temperature was changed within the physiological range. These results indicate that the nitrogenase activity in Synechococcus RF-1 is controlled by a "circadian clock" (Huang et al., 1990), a phenomenon which does not occur with other nitrogen-fixing microorganisms.
The nucleotide sequence of the nifHDK operon in heterocystous cyanobacteria has been well characterized. However, there have been no reports on the molecular structure of the nif genes in unicellular cyanobacteria. In this report, the nucleotide sequence of nifHDK operon in Synechococcus RF-1 is examined.
Materials and Methods
Isolation of DNA from Synechococcus RF-1
Axenic Synechococcus RF-1 were grown in BG-110 according to the conditions described previously (Chou et al., 1989). Since the surface of Synechococcus RF-1 contained an unknown compound which will interact with lysozyme, the cells collected by centrifugation were washed twice in distilled H2O by vibration with Vortex (Model K-550-G) at the maximum speed for 1 min at room temperature. The cells were transferred to 5 ml 6 M guanidine-HCl and vibrated for 1 min (Votex) at room temperature. The guanidine-HCl treatment was repeated once. The cells were then washed with distilled H2O three times to remove the guanidine-HCl and then suspended in 10 mM Tris-EDTA buffer (TE), pH 8.0. Lysozyme was added to have a final concentration of 10 mg/ml. The cells were then in
3Corresponding author. Fax: 886-2-7827954.