Bot. Bull. Acad. Sin. (1996) 37: 173-180
Gang and Weber Genetic variability in wheat smut (Tilletia)
Using random amplified polymorphic DNA to analyze the genetic relationships and variability among three species of wheat smut (Tilletia)
David R. Gang1 and Darrell J. Weber2
Department of Botany and Range Science, Brigham Young University, Provo, Utah 84602, USA
(Received November 17, 1995; Accepted April 9, 1996)
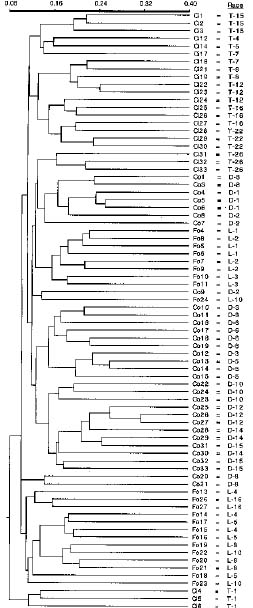
Abstract. Covered smut or bunt of wheat, caused by Tilletia caries, T. foetida (T. laevis), and dwarf bunt, caused by T. controversa, are significant pathogens of wheat. However, the molecular and genetic make-ups of these fungi have not been studied extensively. The extremely long time required to germinate the Tilletia controversa teliospores, their thick walls and high lipid content have discouraged extensive genetic analyses of the fungi by methods such as RFLP or RAPD. We recently developed a method to extract genomic DNA from the thick-walled teliospores of Tilletia. This method was used to extract the genomic DNA from 90 smut samples. The average Tilletia DNA yield per infected wheat head was significantly different for T. controversa as compared to T. caries and T. foetida (T. laevis). RAPD reactions were performed with the 90 Tilletia individuals and 13 random primers. The resulting RAPD patterns were scored for presence or absence of specific fragments and this information was tabulated for each sample in a large data matrix. The total number of RAPD fragments per individual and per primer were calculated. A high level of genetic variability was observed between species, races, and even individuals of the same race of Tilletia. Cluster analysis of the RAPD banding patterns distinguished races of Tilletia caries, T. foetida (T. laevis), and T. controversa. No race- or species-specific markers were identified.
Keywords: Genetic analysis; Genomic DNA; Random Amplified Polymorphic DNA; Tilletia.
Introduction
Covered smut or bunt of wheat is present in all wheat-growing areas of the world. Smut infection has caused considerable economic loss (Grey et al., 1986; Trione, 1982). Common bunt is caused by Tilletia caries (DC) Tul. and T. foetida (T. laevis) (Wallr.) Liro. Dwarf bunt is caused by T. controversa Kuhn. The teliospores of Tilletia species are the major means of spreading the infection.
Although the disease symptoms of dwarf bunt are distinct from the symptoms of common bunt, it is difficult to distinguish between the teliospores of the two pathogens even at the electron microscope level (Hess and Trione, 1986). Stockwell and Trione (1986) had limited success distinguishing between two of the Tilletia species using fluorescence microscopy. Aside from the highly impractical method of infecting wheat kernels with teliospores and waiting for the disease symptoms to appear, a method to definitively distinguish between the species and races is yet to be found.
One method that may provide a means to distinguish between the species of Tilletia is Random Amplified Polymorphic DNA (RAPD) analysis. This method has been
used to distinguish populations of plants by their DNA fragments (Gang and Weber, 1995a; Yu and Pauls, 1993). Jones and Dunkle (1993) used RAPD analysis of mycelium to distinguish races of Cochliobolus carbonum. Strongman and MacKay (1993) were able to distinguish between two subspecies of Hirsutella longicolla (var longicolla and var cornuta) using RAPD analysis of the mycelium. RAPD analysis is also a rapid and efficient method for identifying the degree of genetic similarity and variability among biological populations (Tingey and del Tufo, 1993; Gang and Weber, 1995a).
For many fungal species, however, using the mycelium as a source of DNA for RAPD and other genetic analyses, as was the case in the investigations mentioned above, is impractical. Genetic analysis of Tilletia has been hampered by the extremely long time necessary to germinate the teliospores. Three days are needed to germinate T. caries (Weber and Trione, 1980). Thirty days are required to germinate T. controversa (Trione, 1982). Even though Russell and Mills (1993) have made some progress recently with a cross of T. controversa and T. caries mycelia, large-scale genetic analyses of Tilletia such as with RAPDs, PCR, or RFLPs require a method of extracting DNA from the teliospores. The thick wall, high lipid content (Weber and Trione, 1980), and dormancy of the teliospores have previously prevented the development of such a method. This should no longer remain the case. We recently reported a method that extracts RAPD qual
1Current Address: Institute of Biological Chemistry, Washington State University, Pullman, Washington 99164, USA.
2Corresponding author. Fax: 801-378-7499.