Bot. Bull. Acad. Sin. (1996) 37: 191-196
Chiu et al. N loss in mangrove soil
Nitrogen nutritional status and fate of applied N in mangrove soils
Chih-Yu Chiu1,3, Shui-Cheng Lee2, Hau Tsueng Juang1, Ming-Tan Hur1, and Yuan-Hsun Hwang1
1Institute of Botany, Academia Sinica, Taipei, Taiwan 11529, Republic of China
2Institute of Nuclear Energy Research, Atomic Energy Council, P. O. Box 3 Lungtan, Taiwan, Republic of China
(Received December 19, 1995; Accepted March 19, 1996)
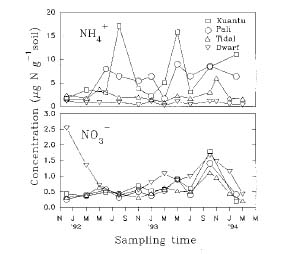
Abstract. Fluctuation of inorganic N content in mangrove soils of the Tamshui estuary, northwestern Taiwan, showed that ammonium concentrations ranged from 0.15 to 17.10 mg N kg-1 soil, while nitrate ranged from trace amounts to 2.54 mg N kg-1 soil. Geographic and edaphic factors caused the difference of inorganic N between the sites. These levels were much higher than values reported elsewhere. Pot experiments showed the added 15N labeled ammonium (6 mg N per pot) disappeared rapidly. The 15N residues remaining in the soil were mostly in the organic form. N uptake by Kandelia candel was 13.1% after one month and 19.6% after three months. Recoveries of applied N after three months were 40.3% in planted treatment and 32.6% in unplanted treatment. Most of the N loss, occurring in the first month, can be attributed to denitrification. The large N loss suggests a high potential for mangrove soils to remove high input of N from the river through denitrification.
Keywords: Denitrification; Kandelia candel; Mangrove; 15N.
Introduction
Nitrogen (N) has been indicated as a major factor limiting the growth of halophytes in intertidal areas (Stewart et al., 1979). Higher productivity of various kinds of halophytes seems to be induced by an additional supply of N (Tyler, 1967; Valiela and Teal, 1974). Evidence suggests that mangrove forests are generally nutrient limited with N (Onuf et al., 1977; Boto and Wellington, 1983; Boto and Wellington, 1984). Positive growth responses to added N were found in mangrove Avicennia marina (Boto et al., 1985; Naidoo, 1987). N was also considered a limiting factor for microbial activity in the mangrove swamp of the Indus Delta (Kristensen et al., 1992).
On the other hand, natural and artificial wetlands have been used to control and remove N in contaminated wastewaters discharges (Brodrick et al., 1988). Biological nitrification-denitrification reaction is important as an N removal mechanism in freshwater marshes (Reddy et al., 1989; Lindau and DeLaune, 1991), and in estuarine sediments (Jenkins and Kemp, 1984). It has been suggested that the denitrification in mangrove sediment, together with the assimilation of N by plants, might improve water quality in the eutroficated river (Nedwell, 1975).
However, information is limited about the effect and impact of nutrient enrichment on mangrove ecosystems. Previous research has shown that the mangrove forest of the Tamshui estuary in northwestern Taiwan, is polluted by municipal sewage (Chiu and Chou, 1991). The objectives of this study are to determine the status of N flux in
this polluted mangrove swamp throughout the year and to evaluate the extent of the N loss through denitrification of mangrove soil.
Materials and Methods
Field Site and Sampling
The Tamshui estuary is one of the most important preserves of mangrove in Taiwan. It includes three major areas (Figure 1). Chuwei, the largest site of all, can be divided into two areas, Tidal mangrove and Dwarf mangrove, depending on the elevation and the vegetation. Details of the ecological and edaphic information of Chuwei have been previously discussed (Chiu and Chou, 1991). Pali, located at about 1 km from rivermouth, was the outermost downstream site of all mangroves. A great number of bird nests were found in this area. Kuantu was the mangrove furthest from the shore, closest to the source of polluted sewage from metropolitan Taipei.
Surface soil (0 to 20 cm) samples were periodically collected from those sites from November 1992 to March 1994. The samples were kept in an ice box and brought back to the laboratory for chemical analysis.
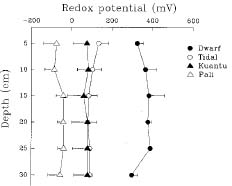
Vertical change of redox potential (Eh) of soils were measured during ebb tide by a portable pH/mV meter (Jenco 6009) with Pt-electrode on May 25 and May 26, 1992. The calibration standard used for Eh was equimolar (M/300) solutions of potassium ferricyanide and potassium ferrocyanide in 0.1 M KCl potassim chloride. The system has an Eh of 0.430 mV at 25°C (Zobell, 1946). Five replicates were measured at each site.
3 Corresponding author.