Bot. Bull. Acad. Sin. (1996) 37: 197-200
Kao et al. Growth of rice cells on ammonium
Polyamines in relation to ammonium-inhibited growth in
suspension-cultured rice cells
Shu Jiuan Chen and Ching Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received January 23, 1996; Accepted March 22, 1996)
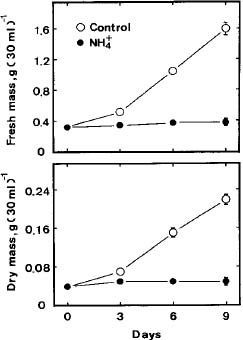
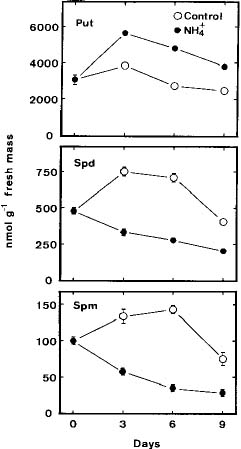
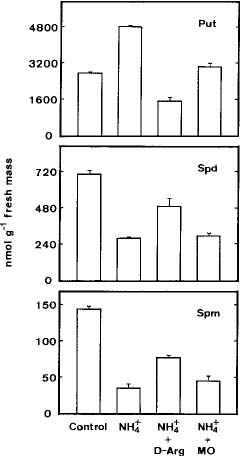
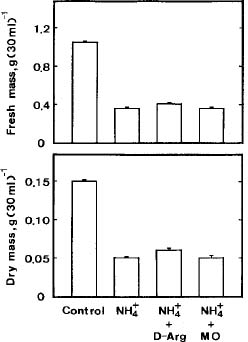
Abstract. The role of endogenous polyamines in affecting the growth inhibition of rice cells fed with ammonium was investigated. When ammonium was used as the sole nitrogen source, the growth of rice cells, as judged by dry or fresh mass, was markedly inhibited. Ammonium-fed rice cells had a higher level of putrescine (Put) than the control cells (supplied with both nitrate and ammonium). However, ammonium-fed cells had much lower levels of spermidine (Spd) and spermine (Spm) than the control. The growth inhibition of rice cells induced by ammonium could not be recovered by the addition of Spd and Spm. D-Arginine or a-methylornithine, inhibitors of Put biosynthesis, caused a reduction in the level of Put in ammonium-fed rice cells, but was unable to reverse the growth inhibition of rice cells fed with ammonium. It was concluded that endogenous polyamines might not play a role in influencing the ammonium-inhibited growth of rice cells.
Keywords: Ammonium; Cell growth; Oryza sativa; Putrescine; Spermidine; Spermine.
Introduction
Polyamines occur ubiquitously in animals, plants, and prokaryotes, and their roles in affecting growth, development, and stress metabolism have been reviewed (Evans and Malmberg, 1989). In both animal cells and bacteria, changes in polyamine levels have been shown to accompany growth (Herbst and Snell, 1948; Tabor et al., 1982; Cohen et al., 1984). Polyamine levels and plant growth rates are positively correlated under a wide variety of conditions in which high levels of polyamines are associated with rapidly growing tissues (Evans and Malmberg, 1989). In contrast, Ozawa and Tusjii (1993) demonstrated that soybean plants accumulated sufficient putrescine (Put) and spermidine (Spd) in their nodules to inhibit the growth of bacteroids of Bradyrhizobium japonicum strain 138NR. In a recent work, Rea et al. (1995) also observed that Spd inhibited growth of maize roots.
Plant cell cultures do not usually grow on ammonium as a sole source of nitrogen (Gamborg and Shyluk, 1970; Behrend and Mateles, 1976; Fukunaga et al., 1978; Kaul and Hoffman, 1993). It has been shown that plants accumulate Put when grown with ammonium instead of nitrate (Hohlt et al., 1970; Klein, 1979; Smith, 1984). Recently, we demonstrated that Put accumulation was a factor causing growth inhibition of rice cells under potassium deficiency (Sung et al., 1994). However, it is not known whether ammonium-inhibited growth of rice cells is also mediated via Put accumulation.
The present investigation was designed to examine the role of endogenous polyamines in affecting the growth inhibition of rice cells fed with ammonium. We present data showing that endogenous polaymines play no role in the control of ammonium-inhibited growth of rice cells.
Materials and Methods
Rice (Oryza sativa cv. Tainan 5) suspension cultures were initiated from calli derived from immature embryo (Yu et al., 1991) and subsequently maintained at 25°C by weekly transfers to fresh liquid medium of Murashige and Skoog (1962) supplemented with 3% sucrose and 5 mM 2,4-dichlorophenoxyacetic acid. Polyamines and inhibitors of Put biosynthesis were sterilized by filtration before adding to the culture medium.
Suspension-cultured cells were collected by filtration through a 400-mesh nylon sieve, and blot-dried on paper towels. The growth of rice cells was measured by fresh mass and dry mass. Dry mass determination was made after drying at 80°C for 48 h. For polyamine determination, the collected cells (about 50 mg fresh mass) were homogenized in 5 ml of 5% perchloric acid. Polyamine levels were determined using high performance liquid chromatography after benzoylation as described previously (Chen and Kao, 1991).
For all measurements, each treatment in an experiment was performed four times, and all experiments were performed three times. Similar results and identical trends were obtained each time; therefore, only one set of data from a single experiment is presented.
1Corresponding author.