Bot. Bull. Acad. Sin. (1996) 37: 201-207
Lee and Lin Peroxidase activities in rice roots
Peroxidase activity in ethylene-, ABA-, or MeJA-treated rice (Oryza sativa L.) roots
Tse-Min Lee1 and Yaw-Huei Lin2
Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 115, Republic of China
(Received January 30, 1996; Accepted May 3, 1996)
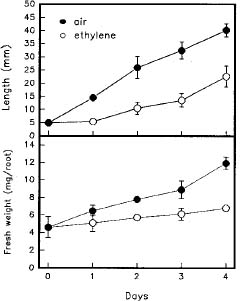
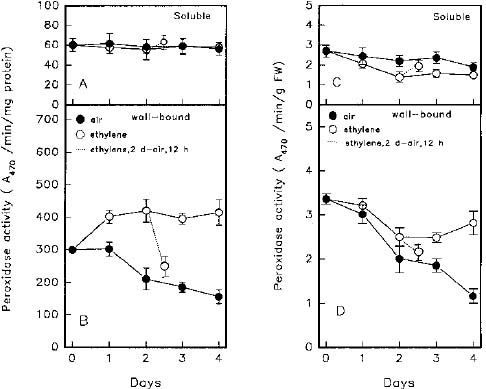
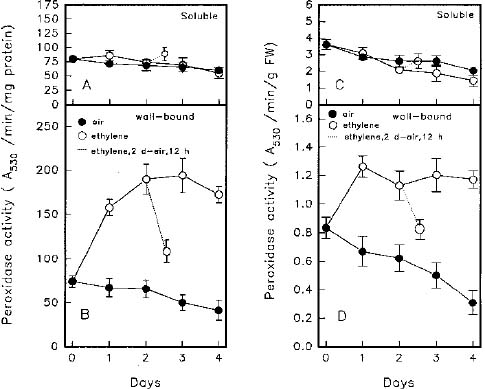
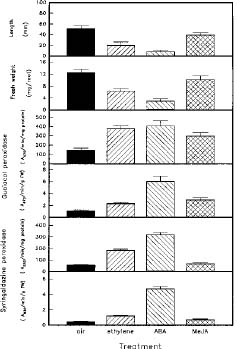
Abstract. Changes of peroxidase (EC 1.11.1.7) activity in ethylene-, ABA- or methyl jasmonate (MeJA)-treated rice (Oryza sativa L. cv. Taichung Native 1) roots were investigated. Guaiacol and syringaldazine were used as substrates for peroxidase activity determination. On a protein or fresh weight basis, activity of wall-bound guaiacol and syringaldazine peroxidase in air-grown roots decreased as growth advanced. In air-grown roots, soluble peroxidase activity on a fresh weight basis also decreased, but activity on a protein basis remained constant. In response to 10 ml/l ethylene, root growth was strongly inhibited while, in contrast, activity of cell wall-bound guaiacol and syringaldazine peroxidase on a protein or fresh weight basis increased. Activity of soluble peroxidase on a fresh weight basis in ethylene-treated roots also increased, although soluble peroxidase activity per mg protein did not change. ABA (10 mM) or MeJA (50 mM) also resulted in an inhibition of root growth but an increase of wall-bound guaiacol and syringaldazine peroxidase activity. Thus, activity of wall-bound peroxidase is inversely related to plant hormone-mediated growth inhibition of rice roots.
Keywords: Oryza sativa L.; Root; Ethylene; ABA; Methyl jasmonate; Cell wall-bound peroxidase.
Abbreviations: ABA, abscisic acid; FW, fresh weight; MeJA, methyl jasmonate.
Introduction
It is known that peroxidase (EC 1.11.1.7) influences several reactions involved in cell wall formation. The ferulic acid is esterified to the arabinoxylans and the peroxidase is believed to catalyze the formation of the diferuloyl cross-links between arabinoxylan chains. Peroxidase is involved with the assembly of lignin and may be involved in linking protein and lignin in the wall (Fry, 1986; Van Huystee, 1987; Whitmore, 1978) subsequently decreasing wall extensibility (Fry, 1979). In peanut hypocotyls (Zheng and Van Huystee, 1992), mung bean hypocotyl (Goldberg et al., 1987) and dwarf sorghum (Schertz et al., 1971), an elevated peroxidase activity is associated with the decrease of plant growth. Studies on two genotypes of tall fescue (Festuca arundinacea Schreb.) with different lengths of leaf elongation zones also showed that the cessation of elongation is closely related to the increase of wall-bound peroxidase activity (MacAdam et al., 1992). Wall-bound peroxidase is involved in the ethylene-regulated growth of higher plants. In the case of Pharbitis, ethylene, which caused the inversion-induced growth inhibition of shoots, is suggested to stiffen the wall by increasing activity of wall-bound peroxidase (Prasad and Cline, 1987). The ethylene-mediated inhibition of pea
epicotyl elongation is accompanied by increasing of peroxidase activity (Ridge and Osborne, 1970). In Ranunculus sceleratus L., ethylene that promotes petiole growth results in decreasing of peroxidase activity (Horton, 1993).
The growth of rice roots is regulated by several plant growth regulators. Ethylene at high concentrations inhibits root growth in etiolated rice seedlings (Jackson, 1982). ABA inhibits rice root growth (Horton, 1991; Lee et al., 1994) and methyl jasmonate (MeJA) also inhibit the growth of rice seedlings (Yamane et al., 1981),
At present, the metabolic processes involved in plant growth regulator-mediated growth inhibitions of rice roots are still unknown. To obtain further information as to whether peroxidase is associated with growth of rice roots, the activities of soluble and cell wall-bound (NaCl extractable) peroxidase were determined in this study. The H+ donor, guaiacol, was used for peroxidase activity determination. Since syringaldazine is thought to be oxidized by peroxidase bound to cell walls that have undergone lignification (Goldberg et al., 1983), it was also used for peroxidase activity determination.
Materials and Methods
Plant Cultivation and Plant Growth Regulator Treatment
Rice (Oryza sativa L. cv. Taichung Native 1) seeds were sterilized with 5% sodium hypochlorite for 10 min. After
1 Present address: Institute of Marine Biology, National Sun Yat-Sen University, Kaohsiung, Taiwan 804, R.O.C.
2 Corresponding author.