Bot. Bull. Acad. Sin. (1996) 37(4): 255-259
Chen and Kao Ammonium and maize leaf senescence
Ammonium accumulation in relation to senescence of detached maize leaves
Shu Jiuan Chen and Ching Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received June 22, 1996; Accepted August 14, 1996)
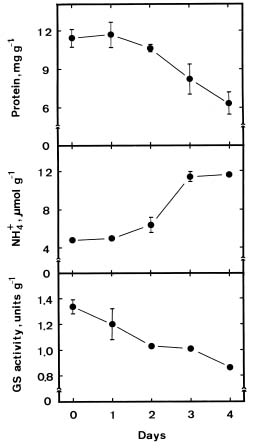
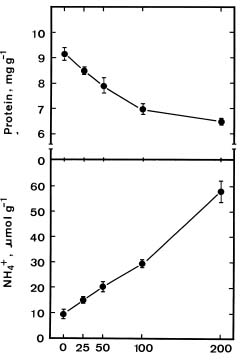
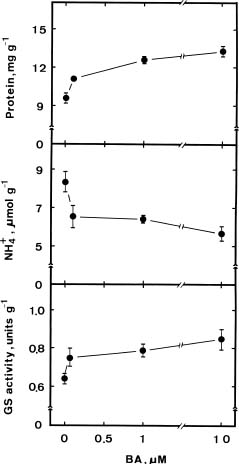
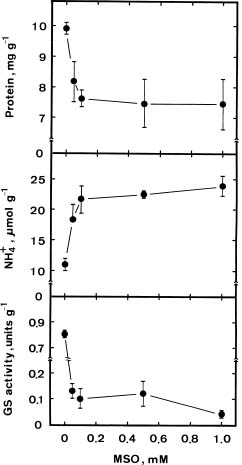
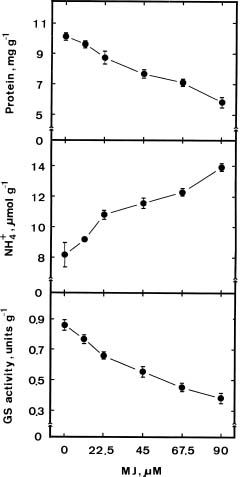
Abstract. We investigated the role of ammonium in the regulation of dark-induced senescence of detached maize leaves. Ammonium levels were found to increase prior to senescence. The accumulation of ammonium was associated with a decrease in glutamine synthetase (GS). Exogenous NH4Cl and methionine sulfoximine (MSO), an inhibitor of GS, increased ammonium level and promoted senescence. Benzyladenine, a synthetic cytokinin, retarded senescence and decreased ammonium level. Methyl jasmonate promoted senescence and increased the ammonium level. The addition of L-glutamine had no effect on dark-induced, MSO- and MJ-promoted senescence. Our results suggest that an increase in ammonium level is associated with the senescence of detached maize leaves in the dark.
Keywords: Ammonium; Benzyladenine; Leaf senescence; Methyl jasmonate; Methionine sulfoximine; Zea mays.
Introduction
Glutamine synthetase (GS, EC 6.3.1.2) is the primary enzyme responsible for ammonium assimilation in plants (Lea and Miflin, 1974). It has been shown that GS activity in leaves decreases during senescence (Kar and Feierabend, 1984; Peeters and Van Laere, 1992; Postius and Jacobi, 1976; Simpson and Dalling, 1981; Storey and Beevers, 1978; Streit and Feller, 1983). Decreased GS activity during leaf senescence may result, at least in part, in the accumulation of ammonium in leaves. In fact, the accumulation of ammonium in senescing leaves has already been described (Postius and Jacobi, 1976; Thomas, 1978). Ammonium is thought to be toxic to plant cells (Givan, 1979) and it has been suggested that ammonium accumulation may be a factor contributing to loss of quality of harvested aspargus spears or leaf senescence (Hurst et al., 1993). However, no clear evidence has been provided to prove this suggestion. Ammonium has been shown to accumulate after the onset of senescence in detached wheat leaves (Peeters and Van Laere, 1992; Thomas, 1978). In the present investigation, we used detached maize leaves to determine whether ammonium accumulation is associated with leaf senescence.
Materials and Methods
Plant Materials and Incubation Conditions
Seedlings of maize (Zea mays cv. XL 678) were grown in vermiculite in a greenhouse with natural light at 30°C day/25°C night for 7 days, by which time the primary leaves were fully expanded. The apical 2.5-cm segments
were excised from the primary leaves and then floated in a glass Petri dish containing 10 ml of distilled water or test solution. Incubation was carried out at 27°C in the dark.
Determination of Protein and Ammonium
For protein determination, leaf segments were homogenized in 50 mM sodium phosphate buffer (pH 7.5). The extracts were centrifuged at 17,600 g for 20 min, and the supernatant liquids were used for determination of protein by the method of Bradford (1976). Protein level was expressed as mg g-1 fresh weight. For ammonium determination, leaf segments were homogenized in 0.3 mM sulphuric acid (pH 3.5). The homogenate was centrifuged for 10 min at 39,000 g, and the supernatant liquids were used for determination of ammonium by the method described previously (Lin and Kao, 1996). Ammonium levels were expressed as µmol g-1 fresh weight.
GS Assay
Leaf segments were homogenized with 10 mM Tris-HCl buffer (pH 7.6, containing 1 mM MgCl2, 1 mM EDTA and 1 mM 2-mercaptoethanol) in a chilled mortar and pestle. The homogenate was centrifuged at 15,000 g for 30 min, and the resulting supernatant was used for determination of GS activity. The whole extraction procedure was carried out at 4°C. GS was assayed by the method of Oaks et al. (1980). The reaction mixture contained in a final volume of 1 ml: 80 µmol Tris-HCl buffer, 40 µmol L-glutamic acid, 8 µmol ATP, 24 µmol MgSO4, and 16 µmol NH2OH; the final pH was 8.0. The reaction was initiated by the addition of the enzyme extract and after incubation for 30 min at 30°C was stopped by adding 2 ml 2.5 % (w/v) FeCl3 and 5 % (w/v) trichloroacetic acid
1Corresponding author. Fax: 02-3620879.