Bot. Bull. Acad. Sin. (1996) 37(4): 261-264
Dai et al. Polysome translation of heat shock proteins
In vitro polysome translation analysis of heat shock proteins in higher plants
Hwa Dai1,4, Yih-Shan Lo1, Yaw-Huei Lin1 , Manfred Ruddat2 and Kwen-Sheng Chiang3
1Institute of Botany, Academia Sinica, Taipei, Taiwan 11529, Republic of China
2 Department of Ecology and Evolution and 3 Department of Molecular Genetics and Cell Biology, University of Chicago, Chicago, Ill. 60637, USA
(Received April 10, 1996; Accepted September 19, 1996)
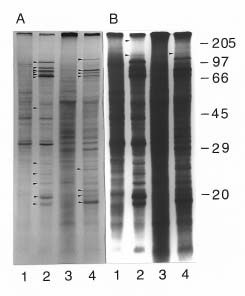
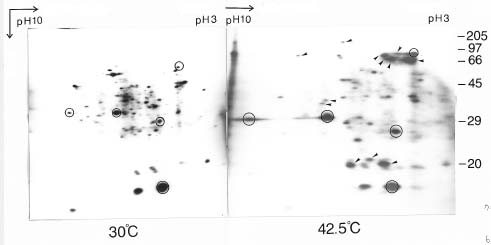
Abstract. We used a simple and highly reproducible method for the analysis of heat shock proteins in rice (Oriza sativa), maize (Zea mays), and mung bean (Vigna radiata). Cytosolic polysomes, isolated from as little as 1_4 grams of seedling biomass, were in vitro translated with the wheat germ system. SDS-PAGE or two Dimensional-PAGE produced excellent separation of the translation products. The active, translating mRNA induced by heat shock was shown by this easy and reproducible method.
Keywords: In vitro polysome translation; Heat shock proteins.
Abbreviation: HSP, heat shock proteins.
Introduction
All organisms, from bacteria to humans, respond to high temperature by inducing or enhancing the expression of a set of heat shock protein (HSP) genes (Schlesinger et al., 1982; Lindquist and Craig, 1988). In higher plants HSPs have been extensively studied (Key et al., 1985; Cooper and Ho, 1983; Tseng et al., 1993 and Vierling, 1991). However, the physiological functions of HSPs are still uncertain. For example, it is known that a proline analogue, azetidine, can induce soybean seedlings to synthesize HSPs, including low-molecular-weight HSPs; however, azetidine-treated seedlings, unlike heat-treated ones, do not acquire heat tolerance (Lee et al., 1996). So much work remains to be done in order to understand the physiological and biochemical roles of HSPs. Most of the published studies were carried out by assaying (1) in vivo heat shock protein synthesis by labeling the newly synthesized proteins in vivo followed by SDS-PAGE analysis, or (2) analyzing heat shock induced mRNA by in vitro translation or by hybridization analysis using an established heat shock gene as a probe. The second method has a chance of picking up preexisting mRNA or non-functional mRNAs, rather than the stress-induced functional transcripts.
In this report, we describe the use of in vitro cell-free translation of polysomes to investigate heat shock proteins induced in seedlings of rice, maize, and mung bean. The heat-induced functional mRNAs can be directly shown by
in vitro translation of the isolated polysomes. The same approach has been done previously in mammalian brains and other organs (Cosgrove and Brown, 1983).
Materials and Methods
Seedling Growth and Heat Shock Conditions
Rice (Oryza sativa L, cv. Tainung 67), maize (Zea mays, cv. Tainung 1), and mung bean (Vigna radiata, purchased from a local store) were used in this study. Rice and mung bean seeds were imbibed in flowing water at 28°C for 72 and 24 h, respectively, before planting in vermiculite. Maize grains were planted directly. Rice and mung bean seedlings were grown in darkness at 30°C. Maize seedlings were grown in darkness at 28°C. Ten-day old rice and maize seedlings and four-day old mung bean seedlings were heat shocked at various temperatures, ranging from 30_42.5°C for 1 h. Polysomes were isolated immediately after heat shock treatment. Exposure to 42.5°C for 1 h yielded the highest amount of heat shock proteins in all three species. Therefore, only these results will be presented.
Polysome Isolation
Seedlings were harvested, and the shoots were cut into approximately 0.5 cm sections. All procedures (modified from Blobel and Sabatini, 1971) were carried out at 4°C. Eight grams of rice seedlings and four grams of maize seedlings were each suspended in 30 ml ice-cold Mpe8 buffer (0.2 M Tris-HCl, 0.4 M KCl, 35 mM MgCl2, 25 mM EGTA, 0.1M b-mercaptoethanol, pH 9.0). Likewise, 20 grams of mung bean seedlings were suspended in 15
4Corresponding author. Fax: 886-02-7827954; E-mail: bodaihwa @gate.sinica.edu.tw.