Bot. Bull. Acad. Sin. (1997) 38: 1_11
Besse and Buchanan Thioredoxin-linked plant and animal processes
(Invited review paper)
Thioredoxin-linked plant and animal processes: the new
generation
Isabelle Besse and Bob B. Buchanan1
Department of Microbial and Plant Biology, University of California, Berkeley, CA 94720, USA
Abstract. The renaissance in the study of thioredoxin in heterotrophic cells that began just prior to this decade has provided new insight into the function and significance of this ubiquitous regulatory disulfide protein. Thioredoxin, reduced enzymically with NADPH via NADP-thioredoxin reductase, has been found to regulate a range of biochemical processes in plants as well as animals. In so doing, thioredoxin targets intramolecular disulfide bonds of proteins such as enzyme inhibitors, seed storage proteins and enzymes. As may be seen, the target proteins have not been identified for a number of thioredoxin functions uncovered in complex systems, such as those involving transcription factors or gene inactivation. In a series of ongoing studies stemming from the fundamental work, thioredoxin has emerged as a new tool in technology and medicine. It now seems clear that we stand at a frontier in which the rapidly expanding knowledge of thioredoxin will lead to applications in industry and health.
Keywords: Thioredoxin; Thioredoxin reductase; Redox regulation.
Contents
Introduction 1
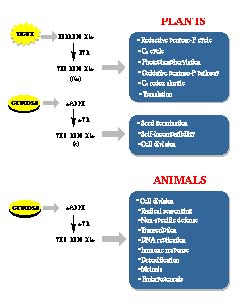
Plant Thioredoxin h 2
Occurrence 2
Proteins Targeted 2
Function 3
New Technologies 3
Animal Thioredoxin 4
Occurrence 4
Proteins Targeted 4
Function 5
Relation to Human Disorders 6
Summary and Perspectives 6
Acknowledgments 7
Literature Cited 7
Introduction
Thioredoxins are 12 kDa proteins with a catalytically active disulfide group that function in a spectrum of biochemical pathways. They have been found in virtually all organisms and were first identified as one of the hydrogen donors for ribonucleotide reductase in Escherichia coli over thirty years ago (Laurent et al., 1964; Holmgren, 1981, 1985). The active site of thioredoxin has two redox-active cysteine residues in a highly conserved sequence [-Trp-Cys-Gly(Ala)-Pro-Cys-]. The oxidized (-S-S-) form of thioredoxin contains a disulfide bridge that is reduced to the sulfhydryl (-SH) level by either reduced
ferredoxin or NADPH via one of two specific enzymes. The reduced form is an excellent catalyst for the reduction of disulfide bonds that are, at best, sluggishingly reduced by glutathione (Holmgren, 1985, 1989; Eklund et al., 1991).
While only one type of thioredoxin has been detected in E. coli or animal cells, three well characterized variants exist in photosynthetic cells. Two of the three (m and f) are located in chloroplasts and can be distinguished from one another on the basis of their primary structure and specificity for target enzymes. The two chloroplast thioredoxins are members of the ferredoxin/thioredoxin system, a regulatory system in oxygenic photosynthesis. Electrons provided by the excitation of chlorophyll are transferred via ferredoxin and an iron-sulfur enzyme, ferredoxin-thioredoxin reductase (FTR) to either of the two
1 Corresponding author. Fax: (510) 642-4995; E-mail: view@ nature.berkeley.edu