Bot. Bull. Acad. Sin. (1997) 38: 29_32
Biswas and Yamauchi Seedling establishment mechanism
Mechanism of seedling establishment of direct-seeded rice (Oryza sativa L.) under lowland conditions
J. K. Biswas1 and M. Yamauchi2
Bangladesh Rice Research Institute (BRRI), Plant Physiology Division, Gazipur-1701, Bangladesh
(Received April 15, 1996; Accepted October 11, 1996)
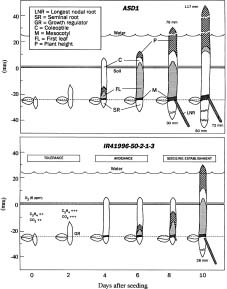
Abstract. The mechanism of seedling establishment under lowland conditions is an important factor for the improvement of direct seeded lowland rice culture. To explore this mechanism, 36-hour-old seedlings of ASD1, IR41996-50-2-1-3, Mahsuri and IR72 were grown in both lowland and drained soil conditions in the IRRI phytotron (29/21°C) for 10 days. The coleoptile tips of the genotypes ASD1 and IR41996-50-2-1-3 were able to penetrate the reduced soil layer to the oxygenated water by 4 days after seeding to allow an O2 transport system to be established. This was followed by the growth of the mesocotyl and first leaf, while the root growth remained inhibited up to 6 days after seeding. ASD1 and IR41996-50-2-1-3 essentially followed the same sequence of growth, but there was an apparent difference in the tolerance of their tissue to oxygen deficit. Mahsuri and IR72 could not elongate their coleoptiles up to the oxygenated water and thus failed to establish prolonged seedling growth under lowland conditions.
Keywords: Anaerobic stress; Coleoptile; Direct seeded rice; Lowland rice; Mesocotyl; Seedling establishment.
Introduction
The adoption of a direct-seeded method for lowland rice culture would significantly decrease the cost of rice production in South East Asia (Flinn and Mandac, 1986). However, at the same time, no varieties have been specifically developed for this purpose. The existing varieties used for lowland culture do not appear to be well-adapted for seedling growth in an initially oxygen-depleted microenvironment. This causes farmers to adopt the costly practice of increasing the seeding rate by 2_3 times. There is therefore an apparent need for varieties which would allow seedling emergence and growth from a soil flooded with a few centimeters of water. The overall pattern of seedling development in rice under conditions of oxygen restriction has been extensively studied (Kordan, 1974a, 1974b, 1975, 1976a, 1976b, 1977a, 1977b; Tsuji, 1972; Turner et al., 1981). Low oxygen conditions severely restrict the growth of roots and leaves on rice seedlings while allowing the growth of the coleoptile and mesocotyl of the shoot. There is some evidence for genotypic variation in this sensitivity to oxygen deprivation (Yamauchi et al., 1993; Turner et al., 1981). While these studies have generated considerable information on the pattern of seedling establishment, very specific data will be required to allow the development and selection of varieties giving a uniformly high rate of establishment
under lowland conditions. This study describes the precise pattern and sequence of growth of four rice varieties in an attempt to evaluate developmental characteristics that would help the establishment of direct-seeded lowland rice.
Materials and Methods
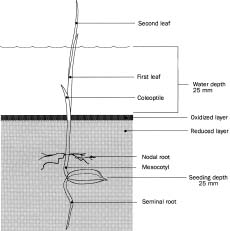
The experiment was conducted in the phytotron (29/21°C) under natural light conditions at the International Rice Research Institute, Los Baños, Philippines in 1993. Two superior genotypes (better seedling establishment under lowland conditions), ASD1 (traditional plant type) and IR41996-50-2-1-3 (modern plant type), and two control genotypes (poor seedling establishment under lowland conditions), Mahsuri (traditional plant type) and IR72 (modern plant type), were grown as a subplot factor of 4 replicated split plot designed experiment. The main plots were assigned for either lowland or drained conditions. Seeds collected from the 1992 harvest and stored at 5°C were used for the study. Table 1 presents the data on seed quality. The germination percentage and germination rate index were calculated as described by Krishnasamy and Seshu (1989) and Yamauchi et al. (1993). Fifteen seeds per unit treatment were soaked with 4 ml of water in a 50 mm plastic petri dish lined with filter paper and allowed to germinate in the dark at 30°C for 36 h. Maahas clay soil of the IRRI farm, passed through a `forty mesh' sieve and mixed with 0.125% corn starch to enrich organic matter, was used for this experiment. Plastic pots 69 mm in diam and 85 mm high were used as the experimental unit. Pots were partially filled to within 25 mm of the top with 190 gm of soil. To simulate lowland conditions, soil
1 Corresponding author. Fax: 88-02-883416 (atten. IRRI Dhaka Office).
2 Present address: Chugoku National Agricultural Experiment Station, Nishifukatsu 6-12-1, Fukuyama, Hiroshima, 721 Japan.