Bot. Bull. Acad. Sin. (1997) 38: 41_44
Ho and Ko Single-spore isolation of fungi
A simple method for obtaining single-spore isolates of fungi
Wang-Ching Ho1,2 and Wen-Hsiung Ko1,3
1Department of Plant Pathology, Beaumont Agricultural Research Center, University of Hawaii at Manoa, Hilo, Hawaii 96720, USA
(Received June 17, 1996; Accepted October 18, 1996)
Abstract. A new method of single spore isolation was developed. A 0.1 µl spore suspension was placed on water agar above one of 50_100 circles marked on the bottom of a plate. After 12_24 h, the number of spores in each circle was counted. Single germinating spores in each circle were transferred separately. For all six species of fungi tested, the number of spores in each micro-drop ranged from 0 to 4. More than 50% of the micro-drops contained a single spore with an unbranched germ tube. This method made it easy to locate well separated spores for single-spore isolation, shortened the isolation time by half, and reduced the incubation period from two days to one.
Keywords: Single-spore isolation; Zoospores; Conidia.
Introduction
Establishing a large number of single-spore isolates is essential for studies of variation, mutation, and segregation in fungi. The conventional method of isolating well separated spores streaked on the agar medium under a stereoscopic microscope (Tuite, 1969) is tedious and time consuming. While studying the mating type change in Phytophthora parasitica Dastur, Ko (1981) developed a relatively easy method for mass isolation of single-zoospore isolates. The method consisted of streaking encysted zoospores on water agar. After incubation at 24°C for 48 h, separated visible colonies were marked and then observed under a light microscope at 100× magnification with 10× objective. Those colonies originating from single zoospores were transferred to V-8 agar plates. This method also has been used to study chemical regulation of mating type in various species of Phytophthora (Ko et al., 1986; Ann and Ko, 1989; Chang and Ko, 1990), to compare the enzyme activity between asexual and sexual progenies of P. parasitica (Ann and Ko, 1990a), to obtain genetic evidence of diploidy for this fungus (Ann and Ko, 1990b), and to document variation in growth rate and colony morphology in P. parasitica induced by exposure to metalaxyl (Chang and Ko, 1992). The method was also used to detect mating type changes during long-term storage of Pythium splendens Braun (Guo and Ko, 1991), to
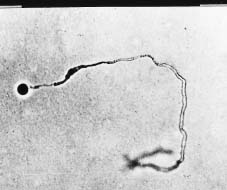
determine whether this fungus was diploid (Guo and Ko, 1994), and to document continuing variation in successive asexual generations of Py. splendens following sexual reproduction (Guo and Ko, 1995). The method developed by Ko (1981) requires time to locate the origin of each colony under the microscope to confirm that the colony originated from a single spore (Figure 1). In this report we describe a simple procedure that eliminates the need to locate the colony origins and, thereby, shortens the time needed to identity single-spore isolates.
2Present address: Department of Plant Protection, National Pingtung Polytechnic Institute, Pingtung, Taiwan, Republic of China.
3Corresponding author. Fax: (808) 969-7923.
Figure 1. A small colony developed from a single zoospore (arrow) of Phytophthora parasitica on water agar 48 h at 24°C (320×).