Bot. Bull. Acad. Sin. (1997) 38: 77-84
Yang et al. Nucleotide sequence of BaMV-V RNA
Nucleotide sequence and phylogenetic analysis of a bamboo mosaic potexvirus isolate from common bamboo (Bambusa vulgaris McClure)
Chi-Chen Yang1,2, Jih-Shiou Liu1, Chan-Pin Lin2 and Na-Sheng Lin1,3
1Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
2Department of Plant Pathology and Entomology, National Taiwan University, Taipei, Taiwan, Republic of China
(Received September 20, 1996; Accepted January 30, 1997)
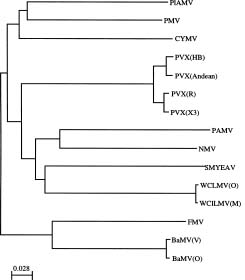
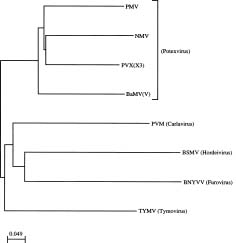
Abstract. The complete cDNA sequence corresponding to the genomic RNA of an isolate bamboo mosaic potexvirus (BaMV-V) from common bamboo (Bambusa vulgaris McClure) was determined. This isolate is the first potexvirus with which a satellite RNA has been associated. The genome organization of BaMV-V, similar to those of other potexviruses, contained five open reading frames (ORFs 1_5) coding for polypeptides with molecular weight of 156 kDa, 28 kDa, 13 kDa, 6 kDa, and 25 kDa, respectively. Nucleotide sequence analysis showed a 10.0% difference from the BaMV-O isolate previously described whereas the amino acid comparison showed a difference of 3.2%. When three conservative domains of RNA dependent RNA polymerase (RdRp) were used for phylogenetic analysis, the greatest variation between two strains of each virus was only 12.8% of that between the two closest members of the potexvirus group. The grouping of potexviruses distinct from other groups of plant viruses was also confirmed by a comparision of three conservative motifs of RdRp.
Keywords: Bamboo mosaic virus; Nucleotide sequence; Potexvirus.
Introduction
Bamboo mosaic virus (BaMV), a member of the potexvirus group, has a flexuous rod-shaped morphology of 490 nm in length and contains a single-stranded positive-sense RNA genome (Lin et al., 1977; 1992; 1994). The complete nucleotide sequence of BaMV from green bamboo (Bambusa oldhamii Munro) (isolate BaMV-O) has been reported (Lin et al., 1994). The RNA genome of BaMV-O is 6366 nucleotides long (excluding poly (A) tail) and contains six putative open reading frames (ORFs 1_6) coding for polypeptides with molecular weights of 155 kDa, 28 kDa, 13 kDa, 6 kDa, 25 kDa, and 14 kDa, respectively. The ORFs 1_5 are common in potexviruses while ORF 6 is divergent.
The ORF 1 encodes RNA dependent RNA polymerase (RdRp), which contains three conservative domainsmethyltransferase, nucleotide-binding, and polymerasecommon in positive strand RNA viruses (Koonin and Dolja, 1993). ORFs 2_4 constitute the triple gene block and are involved in movement of the virus (Beck et al., 1991; Angell et al., 1996). Triple gene block occurs not only in potexviruses but also in carlaviruses (e.g. potato
virus M, PVM) (Zavriev et al., 1991), hordeivirus (e.g. barley stripe mosaic virus, BSMV) (Gustafson and Armour, 1986), furovirus (e.g. beet necrotic yellow vein virus, BNYVV) (Bouzoubaa et al., 1986) and Nicotiana velutina mosaic virus (NVMV) (Randles and Rohde, 1990). ORF 5 encodes the capsid protein of the virus. An ORF 6 has been proposed within the coding region of ORF 1 in papaya mosaic virus (PMV) (Sit et al., 1989), clover yellow mosaic virus (CYMV) (Sit et al., 1990), foxtail mosaic virus (FMV) (Bancroft et al., 1991), potato aucuba mosaic virus (PAMV) (Xu et al., 1994) and BaMV-O (Lin et al., 1994) whereas in narcissus mosaic virus (NMV) (Zuidema et al., 1989), white clover mosaic virus (WClMV) (Beck et al., 1990) and strawberry mild yellow edge-associated virus (SMYEAV) (Jelkmann et al., 1992) an ORF 6 is proposed within or downstream of the coding region of the capsid protein gene. However, the function of the putative ORF 6 remains unknown.
Within the potexvirus group, the overall nucleotide sequence difference among strains of potato virus X (PVX) ranges from 3.7% between X3 (Huisman et al., 1988) and Russian (Skryabin et al., 1988a) strains to 22.6% between Russian and Andean (Orman et al., 1990) strains. On the other hand, the nucleotide sequences of the two strains of WClMV, strain M (Forster et al., 1988) and strain O (Beck et al., 1990), have a 12% divergency. The overall differences in their encoded proteins range from 1.8% to 11.2% among strains of PVX and 4.4% in the two strains of WClMV.
3Corresponding author. Fax: 886-2-782-7954; E-mail: bo124lin@ccvax.sinica.edu.tw
4Sequence data reported from this article have been deposited to the EMBL/GenBank Data Libraries under Accession No. L77962.