Bot. Bull. Acad. Sin. (1997) 38: 85-89
Hung and Kao Senescence of rice leaves
Senescence of rice leaves XXXV. Promotive effects of jasmonates
Kuo Tung Hung and Ching Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received May 29, 1996; Accepted January 18, 1997)
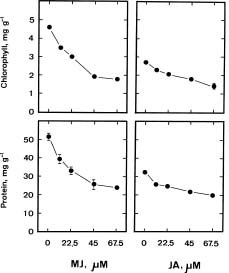
Abstract. Promotion of the senescence of detached rice leaves by jasmonates was investigated. The senescence of detached rice leaves was promoted by linolenic acid, the precursor of the biosynthesis of jasmonic acid, and retarded by inhibitors of lipoxygenase, the first enzyme in the biosynthetic pathway of jasmonic acid. Linolenic acid-promoted senescence was found to be inhibited by lipoxygenase inhibitors. Among the four major lipoxygenase pathway metabolites studied, jasmonic acid and methyl jasmonate were effective in promoting the senescence of detached rice leaves while traumatic acid and trans-2-hexenal had no or only marginal effect on it. The results support a role for endogenous jasmonates in regulating the senescence of detached rice leaves.
Keywords: Jasmonic acid; Leaf senescence; Methyl jasmonate; Oryza sativa L.
Introduction
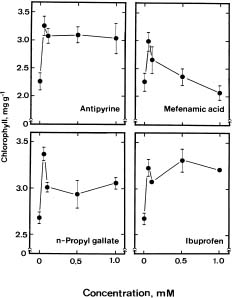
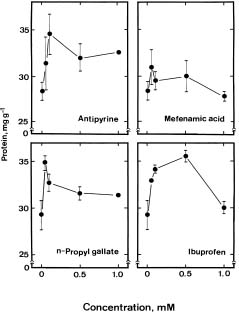
Jasmonates are endogenous substances that have been identified in many plants (Meyer et al., 1984). It has been shown that exogenous jasmonates promote leaf senescence (Chou and Kao, 1992; Parthier, 1991; Ueda et al., 1981; Weidhase et al., 1987). However, it remains to be established whether endogenous jasmonates regulate leaf senescence. In the present investigation, we examined the role of endogenous jasmonates in detached rice leaf senescence by using precursors, several inhibitors of lipoxygenase, the first enzyme in the proposed biosynthetic pathway of jasmonic acid (JA), and major lipoxygenase pathway metabolites including JA, methyl jasmonate (MJ), traumatic acid, and trans-2-hexenal. Both JA and MJ are derived from linolenic acid (18:3) via the lipoxygenase pathway (Vick and Zimmerman, 1984). Trans-2-hexenal and traumatic acid may also be formed from octadecanoid fatty acid linolenic acid by sequential enzyme steps involving lipoxygenase (Siedow, 1991; Vick and Zimmerman, 1984; Zimmerman and Coudron, 1979). The availability of linolenic acid, JA, MJ, trans-2-hexenal, traumatic acid, and several inhibitors of lipoxygenase provides the opportunity to investigate whether endogenous jasmonates regulate the senescence of detached rice leaves.
Materials and Methods
Rice (Oryza sativa cv. Taichung Native 1) was cultured as previously described (Kao, 1980). The apical 3-cm segments excised from the third leaves of 12-day-old seedlings were used. A group of 10 segments, weighing about 45 mg, were floated in a Petri dish containing 10 mL of test solution. Incubation was carried out at 27°C in darkness.
Chlorophyll was determined according to Wintermans and De Mots (1965) after extraction in 96% (v/v) ethanol. For protein extraction, leaf segments were homogenized in 50 mM sodium phosphate buffer (pH 6.8). The extracts were centrifuged at 17,000 g for 20 min, and the supernatants were used for determination of protein by the method of Bradford (1976). Protein and chlorophyll levels were expressed as mg g-1 fresh weight.
All experiments were run at least twice. Data are presented as the results of a single experiment typical of the trend seen in the repeated experiments. MJ [(_)-MJ] was a product of Serva. JA[(±)-JA] and other chemicals were purchased from Sigma Chemical Co.
Results and Discussion
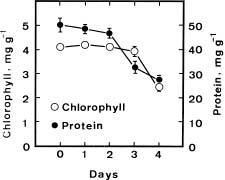
The senescence of detached rice leaves in the dark was followed by measuring the decrease of chlorophyll and protein. Figure 1 shows the time courses of chlorophyll and protein levels of detached rice leaves. Chlorophyll levels remained unchanged for the first 3 days and subsequently declined. A decrease in protein was evident 3 days after leaf detachment.
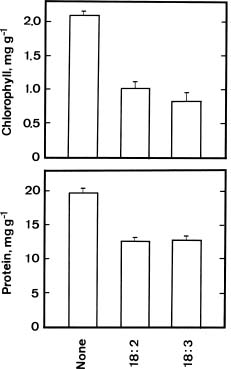
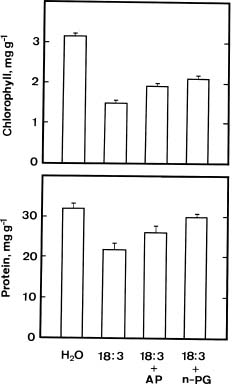
Oxidation of linolenic acid (18:3) by lipoxygenase is the first step in the biosynthesis pathway of JA (Vick and Zimmerman, 1984). The availabililty of linoleic acid, linolenic acid, and several inhibitors of lipoxygenase provided the opportunity to investigate whether endogenous jasmonates regulate the senescence of detached rice leaves. To examine the possible involvement of endogenous JA in regulating senescence of detached rice leaves, we first examined the effects of linoleic acid (18:2), linolenic acid (18:3) and lipoxygenase inhibitors on the senescence of detached rice leaves.
Figure 2 shows the effects of linoleic acid and linolenic acid on the levels of chlorophyll and protein in detached
1Corresponding author. Fax: 02-3620879.