Bot. Bull. Acad. Sin. (1997) 38: 91-95
Das et al. Metabolic changes during rooting in Heritiera
Metabolic changes during rooting in pre-girdled stem cuttings and air-layers of Heritiera
P. Das1, U.C. Basak, and A.B. Das
Mangrove Research Centre, Regional Plant Resource Centre, Bhubaneswar 751 015, Orissa, India
(Received July 3, 1996; Accepted December 2, 1996)
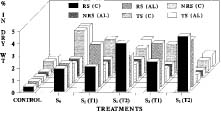
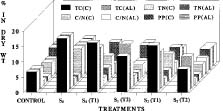
Abstract. The mangroves of the Mahanadi Delta are unique and include 62 of the 64 mangrove species distributed in India. Vegetative propagation through rooting in stem cuttings and air-layers of Heritiera fomes and H. littoralis, commonly known as Sundri, using IAA, IBA, and NAA is reported. Significant increase in the root number was recorded in the air-layers and the pre-girdled stem cuttings of H. fomes treated together with IBA (5,000 ppm) and NAA (2,500 ppm) as compared to H. littoralis. The maximum number of roots was obtained in H. littoralis with IBA (5,000 ppm) and NAA (1,000 ppm). The overall rooting response was better in the treatments with IBA and NAA rather than with IAA and NAA combinations. Variation in the rooting response due to the exogenous application of auxins was also reflected in the metabolic changes during the initiation, emergence, and development of roots in the cuttings and the air-layers. An increase in the level of reducing sugars was noted in the pre-girdled tissues at initiation as well as at subsequent stages of root development, which was further enhanced by the use of auxins. A decrease in the total sugar, carbohydrate, and polyphenols and an increase in the total nitrogen and a high C/N ratio were noted at the root initiation stage in both species. There was less accumulation of photosynthates and nitrogen in the above-girdled tissues in air-layers than the pre-girdled cuttings. Interaction of IBA and NAA promoted starch hydrolysis during root development and subsequently reduced the C/N ratio and increased the protein-nitrogen activity during the development of root primordia. The auxin influenced mobilization of nitrogen to the rooting zone promoted the rooting efficiency in Heritiera.
Keywords: Auxins; C/N ratio; Heritiera; Mangroves; Rooting; Sugars.
Abbreviations: IAA, indole-3-acetic acid; IBA, indole-butyric acid; NAA, a-naphthalene acetic acid.
Introduction
The rich biological diversity of the mangrove ecosystem in the tropical and subtropical inter-tidal regions of the world is an economic resource which has been widely and variously used by coastal people. The mangroves prevent encroachment by the sea by checking soil erosion and thereby stabilizing the shoreline. The organic matter and nutrient flow from the mangrove ecosystem, to a great extent, support the benthic populations of the sea. The Mahanadi Delta is known to have unique communities of plants comprising 62 of the 64 mangrove species distributed in India (Newbold, 1993). Most of these mangroves are threatened, largely by human intervention, resulting in a change in the tidal pattern and salinity gradient. Types of intervention include encroachment of the land for cultivation and shrimp culture, cattle grazing, over exploitation for fuel, timber, and fodder, and others (Das et al., 1994).
Heritiera fomes and Heritiera littoralis, commonly found along the tidal creeks and channels of the coastal swamps (Banerjee, 1990) regenerate naturally through
seeds, but depletion of growing stock, post-dispersal predation of seeds by crabs (Robertson et al., 1990), sporadic flowering, and poor seed set in the remnant mangrove forests necessitate mass vegetative propagation, an alternative to seed propagation, for perpetuation of the species and their re-establishment in the area (Hartmann and Kester, 1989).
The present studies deal with the effect of auxins (IAA, IBA & NAA) on rooting of the pre-girdled stem cuttings and air-layers and the biochemical changes during initiation and development of roots in H. fomes and H. littoralis.
Materials and Methods
One-season-old healthy shoots (semi-hard wood) of Heritiera fomes and H. littoralis, growing in the remnant mangrove forests of the Mahanadi Delta, were girdled 25 cm from the growing tip. After 15 days, the shoots were cut below the girdled point with 2_4 leaves and dipped in tap water. The shoots (2_3 cm from base) were then dipped for 30 sec in IAA, IBA, and NAA solutions alone at 1,000, 2,500, 5,000 ppm concentrations and in combinations of IAA & IBA, NAA & IBA, NAA & IAA at the above concentrations. For air-layering, plant growth regulators in talc were smeared above the girdled portion of the twig,
1Corresponding author. Fax: 91-674-410274.