Bot. Bull. Acad. Sin. (1997) 38: 109-114
Hu et al. Ovule-enriched/specific proteins in Cycas
Identification of ovule-enriched/specific proteins in Cycas taiwaniana using two-dimensional electrophoresis and blotting
Jia-Hwey Hu, Jia-Yu Huang, and Co-Shine Wang1
Agricultural Biotechnology Laboratories, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received August 23, 1996; Accepted January 30, 1997)
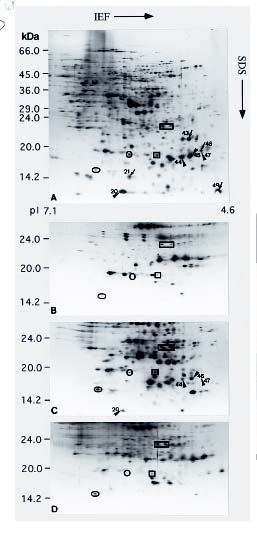
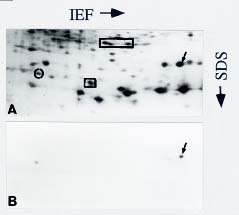
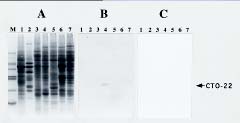
Abstract. We utilized two-dimensional polyacrylamide gel electrophoresis, coupled with silver stain, to identify five ovule-enriched and three ovule-specific polypeptides in Cycas taiwaniana Carr. (CTO). These ovule-enriched and -specific proteins are all with acidic pIs in the 4.8_6.4 range. Polyclonal antiserum to an ovule polypeptide (CTO-22) was prepared and affinity-purified to investigate the specificity of the protein. Immunoblot analyses of total protein from the vegetative organs and from organs of the strobili demonstrated that the CTO-22 polypeptide was ovule-specific. The amino terminal sequence of the CTO-22 protein exhibits no similarity with any known protein. To our knowledge, this is the first ovule-specific protein reported in gymnosperms.
Keywords: Cycas taiwaniana; 2D PAGE; Immunoblotting; Ovule-enriched/specific proteins.
Introduction
Gymnosperms were originally defined by the nature of their female reproductive organs (megastrobili), in which the seeds are naked in the carpel. The gymnosperm ovules are the precursors of seeds that are themselves complex structures. They consist of megasporangia, which except for the micropyle are entirely covered by a number of integument layers (Bold et al., 1987). Although the histological events concerning ovule anatomy and morphology are well characterized both in gymnosperms and in many angiosperms (Johri, 1984; Robinson-Beers et al., 1992), little is known about ovule development at the molecular level. With regard to angiosperms, the relative inaccessibility of the ovule within the ovary and the difficulty of collecting sufficient amounts of ovules have impeded the understanding of the molecular basis of ovule development. Taking advantage of the naked feature and the large size of the ovules, this study attempts to identify ovule-specific proteins in cycads. Previous investigators have identified several mutants affecting ovule and female gametophyte development (Robinson-Beers et al., 1992; Reiser and Fischer, 1993; Léon-Kloosterziel et al., 1994; Modrusan et al., 1994; Gaiser et al., 1995; Angenent and Colombo, 1996). Peroxidases and glycoproteins have been reported to be associated with developing ovules (Pettitt, 1977; Mellon and Triplett, 1989). Nadeau et al. (1996) recently characterized several ovule-specific cDNAs in orchid plants.
Two-dimensional polyacrylamide gel electrophoresis (2D PAGE) coupled with silver stain has been proved to be a sensitive technique for analyzing proteins of mutants
(Santoni et al., 1994), of organs at various developmental stages (Schrauwen et al., 1990; Wang et al., 1992a), and of organs responding to a variety of stresses (Hurkman and Tanaka, 1986; Vartanian et al., 1987). We report herein on the five ovule-enriched and three ovule-specific proteins in Cycas taiwaniana identified by 2D-PAGE analysis. In combination with the use of affinity-purified antibodies, we confirm CTO-22 to be an ovule-specific protein.
Materials and Methods
Plant Materials
Plants of Cycas taiwaniana Carr. were grown in the field. Maturing ovules with weights ranging from 0.5 to 0.6 gm were harvested. Microsporophylls and pollen grains at shedding stage were collected. Leaves (>10 cm) from a newborn leaf crown, entire roots (approximately 12 cm from the apex), and stems were collected separately. Material from both vegetative and reproductive organs (strobili) was dissected and frozen immediately in liquid nitrogen. All material was stored at -80°C.
Preparation, Electrophoresis, and Immunoblotting of Total Protein
Total protein was extracted according to Wang et al. (1992a). Proteins in the phenol phase were precipitated with five volumes of 0.1 M ammonium acetate in methanol at -20°C overnight. The precipitate was washed three times with 0.1 M ammonium acetate in methanol and once with 100% acetone. The pellet was dried and then resuspended in the solubilization buffer consisting of 9.5 M urea, 2% (v/v) Triton X-100, 5 mM K2CO3, 0.5% dithiothreitol (DTT), 500 µgml-1 L-lysine, 0.4%
1Corresponding author. Fax: 886-4-286-1905.