Bot. Bull. Acad. Sin. (1997) 38: 121-129
Chiang et al. Spatial distribution of diatom species composition
Distribution of summer diatom assemblages in and around a local upwelling in the East China Sea northeast of Taiwan
Kuo-Ping Chiang1,4, Fuh-Kwo Shiah2, Gwo-Ching Gong3
1Department of Fishery Science, National Taiwan Ocean University, Keelung, Taiwan, Republic of China
2Institute of Oceanography, National Taiwan University, Taipei, Taiwan, Republic of China
3Department of Oceanography, National Taiwan Ocean University, Keelung, Taiwan, Republic of China
(Received July 30, 1996; Accepted January 18, 1997)
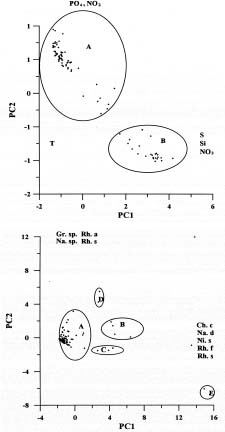
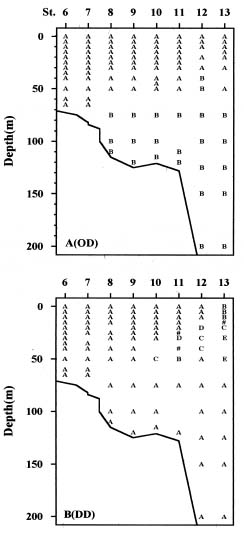
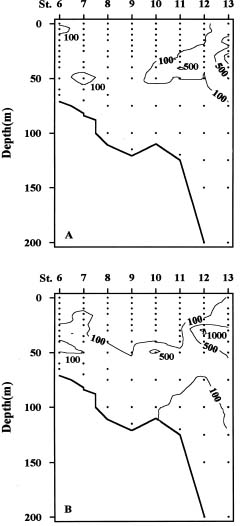
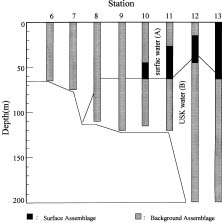
Abstract. Distributions of diatom assemblages in and around a persistent upwelling area off northeastern Taiwan were investigated during summer, 1994. Two water types and two diatom assemblages were defined by principal component analyses. The two water types represent the nutrient-depleted surface water and the nutrient-laden upwelled subsurface Kuroshio Water. During the period of this study, the latter water type did not outcrop but stayed in the subsurface layer, probably resulting from the outflowing of the warm nutrient-poor Taiwan Strait Water. Diatom assemblages indicated the interplay between the two water types. One assemblage, composed of cosmopolitan species in low density, represented the Background Assemblage, which was widely distributed in most of the surface water and also in the underlying upwelling water. The other assemblage, which was observed in the surface water in contact with the underlying upwelling water over the shelf break, represented an Enhanced Assemblage. Some species existing in the Background Assemblage were likely to be enhanced when they were brought into contact with the nutrient-rich upwelled subsurface Kuroshio Water and, subsequently, formed a new assemblage. When the water parcel containing the new assemblage flowed away from the nutrient-rich water, this Enhanced Assemblage sometimes switched back into the Background Assemblage a few days later.
Keywords: Background Assemblage; Diatom assemblage; East China Sea; Kuroshio; Upwelling.
Introduction
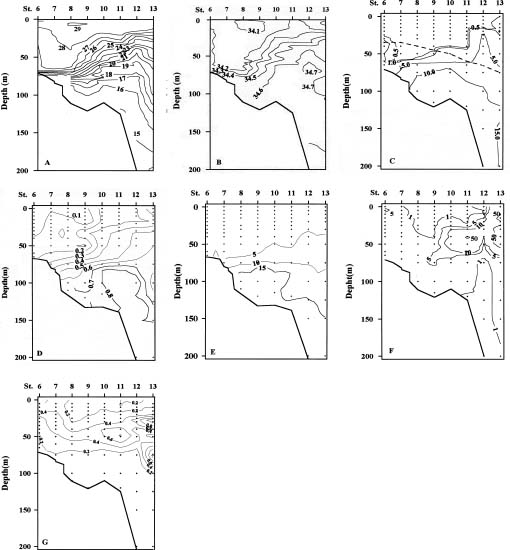
The Kuroshio Current originates from the sea east of the Philippines and flows northward along the east coast of Taiwan. When reaching the area off northeastern Taiwan, it is blocked by the East China Sea shelf and generates a cyclonic cold eddy over the shelf break (Chern and Wang, 1990; Tang, unpublished data). The cold water originates from upwelling of the Kuroshio subsurface Water (Liu et al., 1992a) provided by a counter-current over the continental slope landward of the impinging Kuroshio main stream (Chuang et al., 1993). This cold upwelling has been shown to persist throughout the year (Liu et al., 1992b) and constitutes one of the major nutrient inputs to the East China Sea (Wong et al., 1991; Gong et al., 1995). Outcropping of this upwelling water may be suppressed temporarily by the outflow of Taiwan Strait Water driven by the persistent southwesterly wind in summer (Gong et al., 1992). The residence time of water within the cold eddy has been estimated to be about a week (Liu et al., 1992b).
Light, temperature, nutrient supply, and zooplankton grazing have been recognized as the four most important factors affecting phytoplankton populations (Parsons et al.,
1984; Cullen et al., 1992). Studies of primary production in the southern East China Sea have been intense (Guo, 1991; Chen, 1992; Shiah et al., 1995, 1996); however, essential information regarding species composition of the phytoplankton assemblage in this area is rare. Guo (1991) reviewed the geographical distribution of the phytoplankton in the Philippine Sea, East China Sea, and the northwestern Pacific Ocean. Chen (1992) described a summer phytoplankton community structure in the upwelling area northeast of Taiwan. Possible controlling mechanisms on the distribution patterns of the diatom assemblage were discussed but not fully analyzed in these studies.
It is well recognized that physical and chemical driving forces in the ocean may have a strong impact on phytoplankton production, the growth rate (Shiah et al., 1995, 1996 and citations therein), and assemblage composition (Chiang and Taniguchi, 1993). The study of the spatial and temporal variability of the phytoplankton assemblage may serve as a useful indicator of the interactions among biological, physical, and chemical processes (Chiang et al., 1994 and citations therein). The complex hydrography observed in the East China Sea northeast of Taiwan suggested that such an area could be an ideal experimental ground to examine how physical and chemical processes affect the distribution patterns of phytoplankton assemblages. The study was performed in summer mainly for hydrographical reasons. As mentioned previously, during
4Corresponding author. Fax: (02) 462-1016.