Bot. Bull. Acad. Sin. (1997) 38: 131-139
Moyer and Huang Effect of crop residues on weeds
Effect of aqueous extracts of crop residues on germination and seedling growth of ten weed species
J. R. Moyer1 and H. C. Huang
Research Centre, Agriculture and Agri-Food Canada, P. O. Box 3000, Lethbridge, Alberta, Canada T1J 4B1
(Received September 16, 1996; Accepted December 10, 1996)
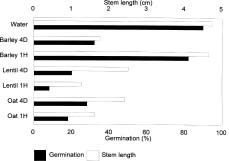
Abstract. Detrimental effects of residues from crops such as canola and lentils on subsequent crops have been observed in petri-dish bioassays and in the field. Suppression of wheat growth by canola and lentil residues has occurred, in producer fields, primarily in the area behind the combine where the residues are concentrated. Adequate straw spreading has permitted producers to grow wheat following canola and lentil crops. The effect of these and other crop residues on common weeds in western Canada has not been assessed. Aqueous extracts of the residues of six different crops were bioassayed for their effect on the germination and seedling growth of ten weeds common in western Canada. Extracts of lentil (Lens culinaris Medic), oat (Avena sativa L.), canola (Brassica napus L.), and barley (Hordeum vulgare L.) were more toxic to flixweed (Descurainia sophia L. Webb), stinkweed (Thlaspi arvense L.), and downy brome (Bromus tectorum L.) than extract of canola was to wheat. The greater toxicity of these crop residues to flixweed, stinkweed, and downy brome than to wheat may permit selective management of these weeds in wheat. Flixweed, stinkweed, and downy brome are major winter annual weeds in winter wheat and usually require late fall or early spring herbicide treatments in no-tillage systems. Therefore, residues of canola, lentil, oat and barley have potential for reducing herbicide use in winter wheat production and in no-tillage direct seeding farming systems. Crop extracts were not toxic enough to affect the growth in the field of seven other weeds in this study.
Keywords: Allelochemical activity; Allelopathy; Bromus tectorium; Canola; Descurainia sophia; Lentil; Oat; Thlaspi arvense.
Introduction
Since the late 1940s producers of agricultural crops have increasingly relied on herbicides for weed control. Problems associated with intensive herbicide use include soil and groundwater contamination, development of herbicide resistant weeds, and the escalating cost of developing new herbicides (Worsham, 1989). Recent assessments of the allelopathic effect of crops or crop residues on weeds has the goal of using naturally produced allelochemicals to reduce reliance on herbicides (Einhellig and Leather, 1988; Putnam and Duke, 1974; Putnam and Duke, 1978).
In the semi-arid plains of western Canada, cereal, oilseed, and pulses are the main annual crops grown. Cultivation techniques that maintain a large percentage of the crop residue on the soil surface to prevent soil erosion by wind have been widely adopted (Moyer et al., 1994). There is a trend in recent years toward zero- or minimum- tillage direct seeding cropping systems that leave nearly all crop residue on the soil surface. Crop residues on the soil surface are positioned such that allelochemicals released by rain are close to the site of weed seed germination (Putnam, 1994; Teasdale et al., 1991). Therefore, there is considerable interest in determining the effect of crop residues on weeds in conservation tillage systems.
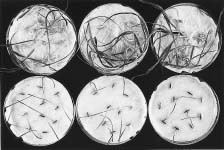
The ability of plants to affect the germination or growth of other plants has been known for centuries (Putnam, 1994). In agricultural crop production the main concern has usually been the effect of toxins from one crop on the yield of the next crop (Guenzi et al., 1967; McCalla and Daley, 1948; Nielsen et al., 1960; Yakle and Cruse, 1984). On the Canadian prairies detrimental effects of toxins from Brassica spp. on the next year's wheat, barley, or flax crops have been reported (Gubbels and Kenaschuk, 1989; Horricks, 1969; Vera et al., 1987). Recent research has aimed at the exploitation of these toxins by developing methods for using them to selectively control weeds in crops (Putnam, 1994). In field experiments it is difficult to test all cropping sequences in the presence of all major weed species to fully understand the effect of crops on weeds. In addition there are potential complex interactions in the field among soil, weather, and allelopathic agents in determining final weed populations. Hence, germination and initial growth of weed seeds in petri-dish tests in the presence of plant extracts have been frequently used to assess the potential toxicity of one plant on another (Hegde and Miller, 1990; Hsu et al., 1989; Martin et al., 1990). Mason-Sedun et al. (1986) found that the toxicity ranking of Brassica spp. on wheat was similar in petri-dish, greenhouse, and field bioassays.
The objective of the research reported in this paper was to assess the effect of residues from major crop species grown on the Canadian prairies on several prevalent weed
1Corresponding Author, LRC Contribution No. 3879664. Fax: (403) 382-3156; Email: MOYER@EM.AGR.CA