Bot. Bull. Acad. Sin. (1997) 38: 153-163
Chiang and Yeh Infectious transcripts of PRSV
Infectivity assays of in vitro and in vivo transcripts of papaya ringspot potyvirus
Chu-Hui Chiang and Shyi-Dong Yeh1
Department of Plant Pathology, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received November 28, 1996; Accepted March 11, 1997)
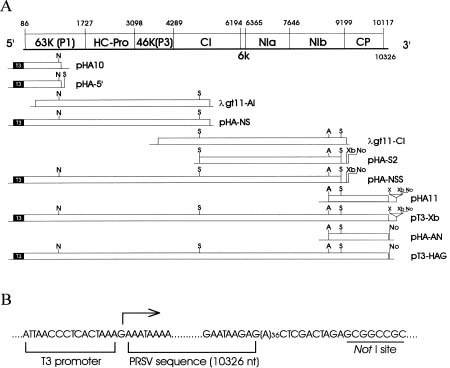
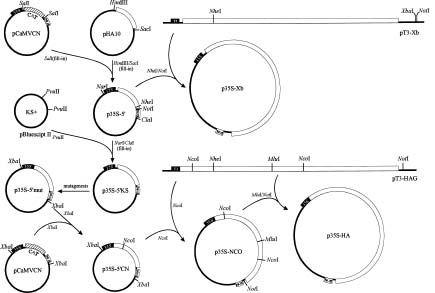
Abstract. A full-length cDNA with nucleotide sequence representing the genomic RNA of a Hawaii strain of papaya ringspot potyvirus (PRSV HA) was constructed downstream from a bacteriophage T3 promoter in an in vitro transcription vector. The plasmid was able to generate an in vitro transcript corresponding to PRSV RNA (10326 nt) with one extra guanosine residue at the 5' terminus and 12 nonviral nucleotides at the 3' end following a poly(A)36 tract. In vitro translation products and immunoprecipitation analysis with the antiserum to PRSV verified correctness of the gene expression of the transcript. When the capped transcript was mechanically introduced to the systemic host papaya and the local lesion host Chenopodium quinoa, typical symptoms of PRSV HA appeared at almost the same time as on those host plants inoculated with native PRSV RNA. Western blotting and serologically specific electron microscopy with PRSV antiserum confirmed the infection. The uncapped in vitro transcript and the transcript with longer nonviral nucleotides (64 nt) at the 3' end were not infectious. The full-length cDNA was also constructed with a cauliflower mosaic virus (CaMV) 35S promoter and a nopaline synthase (NOS) terminator in an in vivo expression vector. Purified plasmids were applied directly onto host plants either mechanically or by bombardment with a particle delivery system to analyze their infectivity. The plasmid without extra nucleotides between the 35S promoter and the 5' end of PRSV sequence and with 10 nonviral residues and a NotI site at its 3' end was infectious, as evident from symptom development, and ELISA, immunodiffusion, and serologically specific electron microscopy analyses with PRSV antiserum. The construct with 33 nonviral nucleotides at the 5' end of the PRSV sequence and more than 64 nucleotides at the 3' end was not infectious. The infectious in vitro and in vivo transcripts derived from the full-length cDNA to PRSV RNA are the longest so far recorded for a plant virus.
Keywords: In vitro transcript; In vivo transcript; Papaya ringspot potyvirus.
Introduction
Papaya ringspot virus (PRSV) is a member of the genus Potyvirus, the largest group among plant viruses (Ward and Shukla, 1991), and is the major limiting factor for papaya production throughout tropical and subtropical areas (Purcifull et al., 1984). PRSV particles are flexuous rods, 700_800 nm in length, and contain a monopartite genome consisting of a positive-sense ssRNA with a 5' genome-linked protein (VPg) (Siaw et al., 1985; Riechmann et al., 1989; Murphy et al., 1990) and a 3' poly(A) tract (Hari et al., 1979).
The complete nucleotide sequences of a severe Hawaii strain PRSV HA and a mosaic Taiwan strain PRSV YK have been determined (Yeh et al., 1992; Wang, 1993). The PRSV genome contains 10326 nucleotides excluding the 3' poly(A) tail and is the longest encountered among the sequenced potyviruses. PRSV RNA encodes a large polyprotein of 381 kDa (Yeh and Gonsalves, 1985; Yeh et al., 1992) which is processed into 8 or 9 final products via three virus-encoded proteinases (P1, HC-Pro, and NIa). Like other potyviruses, the proposed genetic organization
of PRSV RNA is VPg - 5' leader - P1 (63K) - HC Pro - P3 (46K) - CI - P5 (6K) - NIa - NIb - CP - 3' noncoding region - poly(A) tract (Yeh et al., 1992). The P1 protein of PRSV is the most variable protein among potyviruses, and it has an Mr of 63 kDa, which is 18_34 kDa larger than those of the other potyviruses (Yeh et al., 1992).
Infectious transcripts constructed from cDNA clones of plant and animal RNA viruses have been well documented (Boyer and Haenni, 1994). In plant viruses, infectious in vitro RNA transcripts were first successfully prepared for brome mosaic virus (BMV) (Ahlquist and Janda, 1984), and the construction of a directly infectious cDNA clone was first reported for RNA 3 of alfalfa mosaic virus (AlMV) through coinoculation with RNAs 1, 2, and 4 (Dore and Pinck, 1988). In the potyvirus group, infectious in vitro transcripts have been synthesized from full-length cDNA clones with bacterial phage promoters for tobacco vein mottling virus (TVMV) (Domier et al., 1989), plum pox virus (PPV) (Riechmann et al., 1990), zucchini yellow mosaic virus (ZYMV) (Gal-On et al., 1991), and tobacco etch virus (TEV) (Dolja et al., 1992). In vivo infectious clones have been obtained for PPV (Maiss et al., 1992), ZYMV (Gal-On et al., 1995) and potato virus Y (PVY) (Fakhfakh et al., 1996), which were driven by a cauliflower mosaic virus (CaMV) 35S promoter and ter
1Corresponding author. Fax: 886-4-2877585; E-mail: sdyeh@ nchu.edu.tw