Bot. Bull. Acad. Sin. (1997) 38: 165-169
Basra et al. Polyamines and heat-shock response of mung bean
Are polyamines involved in the heat-shock protection of mung bean seedlings?
Ranjit K. Basra1, Amarjit S. Basra1,3, C.P. Malik1,4, and I.S. Grover2
1Department of Botany, Punjab Agricultural University, Ludhiana-141004, India
2Department of Botany, Guru Nanak Dev University, Amritsar-143005, India
(Received June 24, 1996; Accepted March 14, 1997)
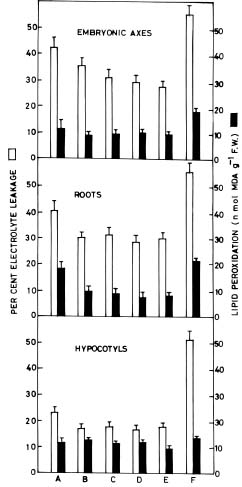
Abstract. Germinating seeds of Vigna radiata (Linn) Wilczek cv. ML 311, with a radicle length of 5 mm, were subjected to a heat-shock episode of 50°C for 2 h followed by transfer to the normal temperature (28°C) for 3 days in the dark. Exogenous effects of polyamines (putrescine, spermidine, and spermine) on the recovery growth and membrane integrity of seedling tissues were studied. Application of polyamines, either as a pre-treatment at 28°C for 2 h prior to heat-shock or as a co-treatment (50°C, 2 h) during the heat-shock period itself, enhanced the recovery growth of both roots and hypocotyls but especially the former with the order of effectiveness being putrescine, spermidine, and spermine. Treatment with polyamine biosynthetic inhibitors, i.e. D,L a-diflouromethylarginine (DFMA) and D,L a-diflouromethylornithine (DFMO) resulted in thermosensitization, making seedlings vulnerable to heat-shock. This effect could be ameliorated by putrescine application. An important role of polyamines in heat-shock protection is thus indicated.
Keywords: Heat-shock; Ion leakage; Lipid peroxidation; Polyamines; Root growth; Seedling growth; Vigna radiata.
Introduction
The diamine putrescine, the triamine spermidine, and the tetramine spermine are ubiquitous in plant tissues and have been implicated in an overwhelming array of plant growth and developmental processes (Bagni, 1989). There is a growing appreciation of the role of polyamines in plant stress responses (Evans and Malmberg, 1989; Flores, 1991), but their role in heat-shock protection of higher plants is not well understood (Basra et al., 1992).
Heat stress is a major factor limiting the productivity and adaptation of crops, especially when it coincides with critical stages of plant growth and development (McWilliams, 1980; Chen et al., 1982; Paulsen, 1994). Even brief episodes of heat-shock between temperature ranges of 45 to 50°C induce marked changes in plant growth processes. The repair of plant cells exposed to heat stress, after having been returned to an optimal temperature, has been reported (Bauer and Senger, 1979), but the degree of recovery and the time required for recovery depend upon the severity of stress (Berry and Björkman, 1980).
In order to have a better understanding of plant responses to heat-shock, it is important to know the regulatory factors involved in imparting protection and growth recovery processes. In the present study, the polyamine-
mediated regulation of the heat-shock response of etiolated mung bean seedlings was investigated using exogenous applications of polyamines and their biosynthetic inhibitors. Changes in electrolyte leakage and lipid peroxidation of seedling tissues were monitored as measures of cellular membrane integrity.
Materials and Methods
Mung bean seeds (Vigna radiata (Linn.) Wilczek cv. ML-131) were germinated in the dark for 24 h at 28°C in Petri-dishes (9.0 cm) on filter paper moistened with 5.0 ml of distilled water. Heat-shock at 50°C for 2 h was applied to germinated seeds with a radicle length of 5 mm followed by transfer to 28°C on water for 72 h in the dark to observe recovery growth. For growth measurements, length of the whole seedling (root plus hypocotyl), primary root, and hypocotyl were measured after the 3 day recovery period.
The tested polyamines were: putrescine, spermidine, and spermine. The polyamine biosynthetic inhibitors were: DFMA (D,L a-diflouromethylarginine) and DFMO (D,L a-diflouromethylornithine). These were applied either as a pre-treatment at the normal temperature of 28°C for 2 h before subjecting the seedlings to heat-shock (50°C, 2 h) or as a co-treatment during the heat-shock period itself. Similarly, lysine, an amino acid, was tested at equivalent concentrations to demonstrate that heat-shock protection is really a polyamine related phenomenon and not simply a growth effect from the extra reduced nitrogen and carbon supplied. After each treatment, the seedlings were
3Corresponding author.
4Current address: Department of Environmental Sciences, Guru Jambeshwar University, Hisar-125001, India.