Bot. Bull. Acad. Sin. (1997) 38: 171-176
Liu et al. Callus formation and endogenous indole-acetic aicd
Effect of hormone treatment on callus formation and endogenous indole-acetic acid and polyamine contents of soybean hypocotyl cultivated in vitro
Zin-Huang Liu1, Wei-Chang Wang, and Sho-Yun Yan
Department of Biology, National Sun Yat-Sen University, Kaohsiung, Taiwan 804, Republic of China
(Received January 14, 1997; Accepted March 28, 1997)
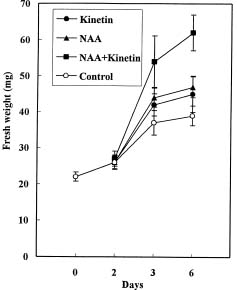
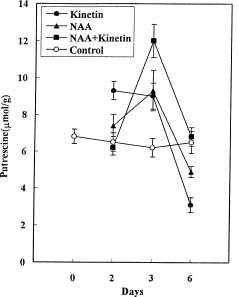
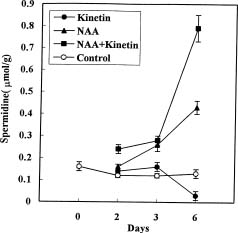
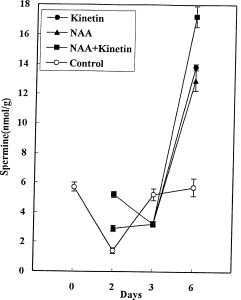
Abstract. Callus was formed and grew well when explants were cultured on medium containing both NAA and kinetin. The endogenous IAA in hypocotyls was accumulated on the medium with both hormones, showing the suppression effect of kinetin on IAA degradation. Besides, the accumulation of NAA in hypocotyl explants was also markedly enhanced by kinetin in the culture medium containing both hormones. In the MS+NAA+Kinetin treatment, the accumulation of spermidine and spermine occurred on the sixth day and was accompanied by rapid callus growth. Increasing putrescine levels were found during the first 3 days in culture when cell multiplication was rapid. The accumulation of endogenous IAA, exogenous NAA, spermidine, and spermine are in proportion to the callus growth in soybean hypocotyl explants.
Keywords: Callus; Glycine max; Indole-3-acetic acid; Polyamines.
Introduction
Cytokinin and auxin are known to promote callus formation in tissue culture (Skoog and Armstrong, 1970; Letham, 1974; Akiyoshi et al., 1983). Auxin has a wide variety of effects on plant growth and morphogenesis. Indole-3-acetic acid (IAA), a natural auxin of higher plants, is involved in regulating cell elongation, cell division and differentiation (Dietz et al., 1990). Cytokinins can promote cell enlargement in certain tissues (Rayle et al., 1982; Ross and Rayle, 1982). Moreover, polyamines, another class of growth regulating substance, are reported to be involved in several aspects of plant development (Berlin, 1981; Galston, 1983; Smith, 1985). Numerous investigations have correlated an increase in polyamine levels with cell division and a drop in polyamines during any subsequent lessening metabolic activity (Heimer and Mizrahi, 1982; Walker et al., 1985). Little is known about the change of endogenous IAA and polyamines affected by the exogenous application of auxin and cytokinin in hormone-treated tissue culture. In the present study, the effect of naphthaleneacetic acid (NAA), a synthetic auxin, and kinetin on the levels of endogenous IAA and polyamines in soybean hypocotyl explants during the formation of callus was investigated.
Materials and Methods
Plant Tissue Culture
Soybean seeds (Glycine max) strain Tk5 were obtained from the Asia Vegetable Research and Development Center in Taiwan. Soybean seeds were collected and washed in tap water followed by one rinse in sterile distilled water. Soybean seeds were surface sterilized for 1 min in 70% ethanol, soaked in 1% sodium hypochlorite plus a drop of a liquid detergent (Tween-20), and then shaken gently (150 rpm) for 20 min, followed by three rinses in sterile distilled water. Seedlings were grown from seeds in culture tubes (diameter 2 cm) containing 10 mL B5 medium (Gamborg et al., 1968). Soybean seedlings were placed in a growth chamber under the following conditions: 27ºC, in darkness, RH 70 to 80% after 7 days, the young seedlings were about 5 cm tall for tissue culture.
Hypocotyl segments (5 mm in length) were cut from the connection of two cotyledons of soybean plants after 7 days of growth in four different media. The basal medium (hormone-free) consisted of inorganic nutrients as found in Murashige-Skoog medium (Murashige and Skoog, 1962) and vitamins of Gamborg's medium, B5 medium, (Gamborg et al., 1968) plus 250 mg/L casein hydrolysate supplemented with 3% sucrose (MS medium). Plant growth regulators, naphthaleneacetic acid (NAA), 2 mg/L, or kinetin, 1 mg/L, or both were added for different treatments (MS+NAA, MS+Kinetin, MS+NAA+ Kinetin). The pH of the medium was adjusted to 5.7 prior to addition of 9% (w/v) agar and sterilized by autoclaving
1Corresponding author. Fax: 886-7-525-3609; Email: zhliu@ mail.nsysu.edu.tw