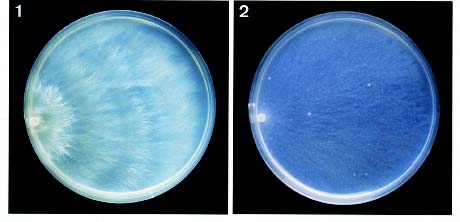
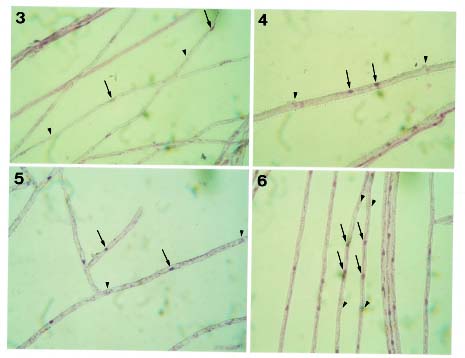
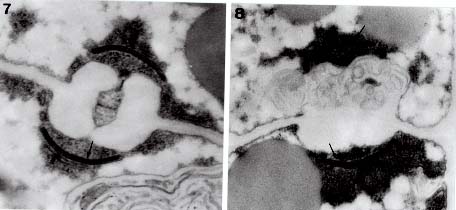
Bot. Bull. Acad. Sin. (1997) 38: 191-195
Wu and Huang Studies of Hyphodontia
Cultural and ultrastructural studies of Hyphodontia mollis and H. subglobosa
Sheng-Hua Wu1 and Yu-Ying Huang
Division of Collection and Research, National Museum of Natural Science, Taichung, Taiwan, Republic of China
(Received December 10, 1996; Accepted March 8, 1997)
Abstract. Cultural and ultrastructural studies are reported for Hyphodontia mollis and H. subglobosa, which are known only from Taiwan. These two species have bipolar sexuality, normal nuclear behavior, and dolipores with continuous parenthesomes. Cultural characters are also provided for both species.
Keywords: Cultural studies; Hyphodontia mollis; H. subglobosa; Taiwan; Ultrastructure.
Introduction
Hyphodontia J. Erikss. is generally regarded by mycologists as a member of the heterogeneous family Corticiaceae Herter, encompassing about 50 species (Langer, 1994). Species of Hyphodontia basically have resupinate basidiomata. Odontioid or grandinioid hymenial surfaces are commonly found in Hyphodontia, but smooth or poroid surfaces are present in some species. The Hyphal system of Hyphodontia is basically monomitic, with nodose-septate generative hyphae present in most species. The Basidia of Hyphodontia are uniformly suburniform and bear fairly small basidiospores, which usually contain one to a few distinct oily drops. The Mycelia of Hyphodontia species generally grow fairly slowly on MEA and usually develop malocysts and/or drepanocysts in cultures (Hassan Kasim and David, 1983). These structures are consistently present in a few species, but typically appear only in some cultures of a species (Nakasone, 1990). Most studied species of Hyphodontia show tetrapolar sexuality and have normal nuclear behavior. Studies of ultrastructure by Langer and Oberwinkler (1993) have shown that Hyphodontia species have dolipores with continuous parenthesomes. This type of parenthesome is not common in the homobasidiomycetes, but has been extensively detected in the heterobasidiomycetes (Wells, 1994).
Most studied species of Hyphodontia show tetrapolar sexuality, with only a few exceptions. Hyphodontia arguta (Fr.) J. Erikss. was reported by Brown (1935) as homothallic. More recently tetrapolarity has been reported for this species (Boidin, 1951; Boidin, 1958; Boidin and Lanquetin, 1965). Hyphodontia efibulata J. Erikss. & Hjortstam was reported by Hassan Kasim and David (1983) as presumed haploid parthenogenetic because uni
nucleate status was indicated for basidiospores, monosporous mycelium, and polysporous mycelium. However, H. efibulata has simple-septate hyphae, whereas most Hyphodontia species bear nodose-septate hyphae. Hyphodontia gossypina (Parmasto) Hjortstam was reported by Hassan Kasim (1981) as having bipolar sexuality. This species was originally known as Fibrodontia gossypina Parmasto. Hjortstam (1990) transferred this species to Hyphodontia, believing the presence of skeletal hyphae insufficient to separate Fibrodontia Parmasto from Hyphodontia. This species (Langer and Oberwinkler, 1993) has dolipores with continuous parenthesomes.
Hyphodontia mollis and H. subglobosa were proposed by Wu (1990) as new species from Taiwan, and both are still known only from Taiwan. At an earlier stage of this study, H. mollis was determined to be bipolar in sexuality. Hyphodontia subglobosa is a species with simple-septate hyphae. As above-mentioned, bipolar sexuality and simple-septate hyphae are rarely present in Hyphodontia. This study was designed to clarify the placement of these two species in Hyphodontia, through the analysis of their sexuality, cultural characters, nuclear behavior, and septal pore ultrastructure.
Materials and Methods
Fungal Specimens and Cultures
Studied fungal specimens and cultures collected from Taiwan, are deposited at the herbarium of the National Museum of Natural Science, ROC (TNM). Monosporous mycelia and polysporous mycelia offered for this study, have been obtained from germinated basidiospores according to Wu (1996).
Cultural Studies
Cultural description and the species code system are basically from Nobles (1965) with amendments by Boidin and Lanquetin (1983). Minor modifications to Nobles'
1Corresponding author. Fax: +886-4-323-5367; Email: shwu@ nmns1.nmns.edu.tw