Bot. Bull. Acad. Sin. (1997) 38: 215-223
Cheng and Chou Ecotypic variation of I. cylindrica in Taiwan
Ecotypic variation of Imperata cylindrica populations in
Taiwan: I. Morphological and molecular evidences
Kur-Ta Cheng2 and Chang-Hung Chou1,2,3
1Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
2Graduate Institute of Botany, National Taiwan University, Taipei, Taiwan, Republic of China
(Received December 9, 1996; Accepted May 3, 1997)
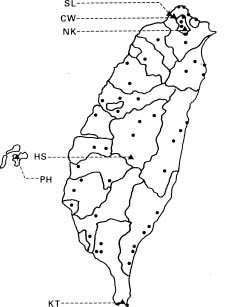
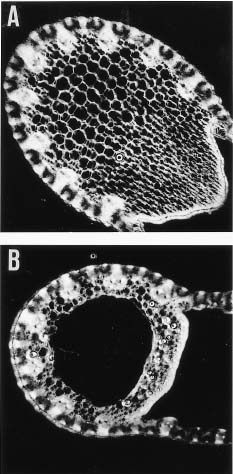
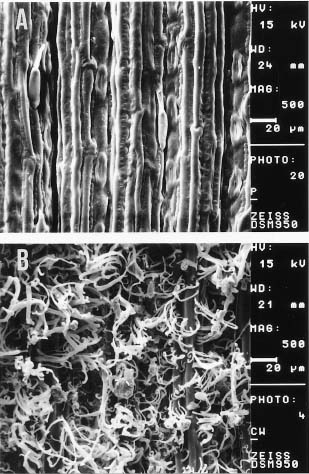
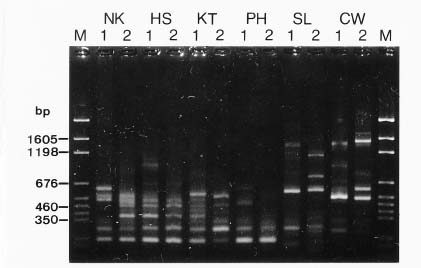
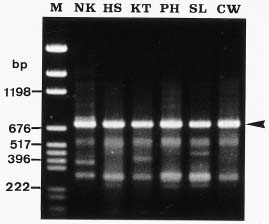
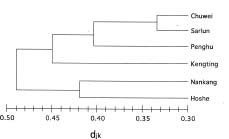
Abstract. Imperata cylindrica L. Beauv, a common C4 grass, is widely distributed in various habitats in Taiwan. Among 55 populations in various habitats, six sites were selected for the study. These sites are namely, Chuwei (an estuary mangrove forest area with high salinity), Sarlun (in a coastal saline area), Kengting (at Taiwan's southern tip with a drought season in winter), Penghu (an islet about 100 km from the west coast that is droughty in most seasons), and Nankang and Hoshe (both inland control sites with mild weather). Leaves of I. cylindrica sampled from these sites were examined under light microscope and scanning electron microscope for anatomic study in order to see their difference. The results showed that the grass leaves collected from the Chuwei site exhibited a clear difference from those of other sites, revealing a hollow structure in the midrib of leaves and villous trichomes on the abaxial leaf surface. The rhizomes of the grass were also collected from each site, brought back to the laboratory, and were transplanted into pots and grown in a greenhouse of the Academia Sinica. The phenotypic characters, such as plant height and leaf width, of the grasses collected from the field and from the greenhouse were compared. The results showed that plants collected from Chuwei, Sarlun, and Penghu were significantly shorter than those of the remaining three sites. The grass leaves collected from both the field and greenhouse were analyzed by means of random amplified polymorphic DNA. Forty 10 base pair primers were employed, and only 31 primers were adequate. Of them, 82 distinguished bands resulted from the RAPD study, showing the difference among populations of the six sites. Furthermore, using an Euclidean distance method, the phylogenetic relationship among the six populations was obtained. The findings revealed that the Nankang and Hoshe populations evolved into an ecotype, which was significantly different from another cluster of the remaining 4 populations. However, populations at Chuwei and Sarlun formed into a unique ecotype although they were also closer to other populations Penghu and Kengting. The phenotypic, morphological, and molecular data are correlated to each other.
Keywords: Cogongrass; Ecotypic variation; Imperata cylindrica; Population; RAPD.
Introduction
Cogongrass (Imperata cylindrica L. Beauv), an aggressive, rhizomatous, and weedy grass, is a troublesome weed in fields throughout the tropical and subtropical regions of the world. Genus Imperata comprises ten species in the world, but only one species is in Taiwan (Hsu, 1975). This grass is widely distributed on the island of Taiwan and its islets, exhibiting its great adaptability to various habitats, particularly in the highly saline coastal areas and drought land. Chou (1989) indicated that I. cylindrica was an allelopathic plant, revealing strong aggressiveness against other plants. It has been listed as one of the seven worst weeds of the world (Holm, 1969). Flint and Patterson (1980) found two ecotypes of Imperata cylindrica (sun ecotype and shade ecotype) in Iraq according to morphological characteristics such as leaf area, leaf weight, leaf/stem ratio, etc. Al-Juboory and Hassawy (1980) collected this grass from fifteen locations throughout Iraq and com
pared their morphological development. They found that plant height and density of stands varied more than two-fold; the number of flowering heads varied eleven-fold among different locations; and mature-leaf width varied from 4.7 to 6.9 mm, reflecting the existence of different ecotypes of I. cylindrica in Iraq. Matumura et al. (1983) and Matumura and Nakajima (1988) described several biotypes of I. cylindrica in Japan based on morphological characters under various environmental stresses, such as drought, salt, etc.
The aforementioned studies emphasized the morphological and phenotypical characters among the ecotypes of I. cylindrica under different habitats and stress environments, yet a biochemical or molecular approach to elucidate the mechanism of the ecotype formation received little attention. A newly developed technique, random amplified polymorphic DNA (RAPD), has recently received high recognition as a powerful and efficient tool with applications to ecology, systematics, evolution studies, and especially to population biology (Halward et al., 1992; Cheng, 1994; Tao et al., 1993; Tinker et al., 1993; Visrling et al., 1994; Welsh and
3Corresponding author.