Bot. Bull. Acad. Sin. (1997) 38: 245-250
Tso and Chen Acid phosphatase isozyme in rice
Isolation and characterization of a group III isozyme of acid phosphatase from rice plants
S. C. Tso and Y. R. Chen1
Department of Botany, National Taiwan University, Taipei, Taiwan, Republic of China
(Received November 15, 1996; Accepted July 18, 1997)
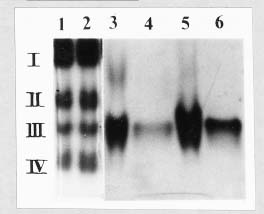
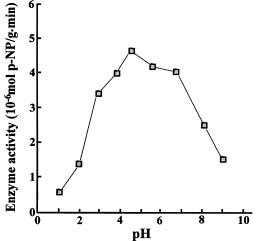
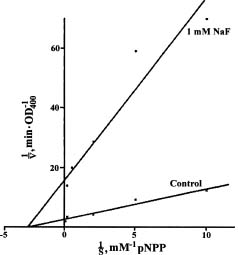
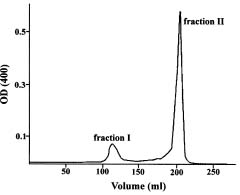
Abstract. The acid phosphatases (EC 3. 1. 3. 2.) of rice seedlings consisted of four groups of isozymes. The group III isozyme was purified through ammonium sulfate, DEAE-Sepharose, Con A-Sepharose, and chromatofocusing. A 198-fold enhancement of activity was attained. This isozyme showed only one band in native-polyacryamide gel electrophoresis. However, gel filtration revealed two peaks of enzyme activity. Their molecular weights were 130 kDa and 100 kDa. The purified isozyme showed a pH optimum of 5. Its Km for p-nitrophenyl phosphate (p-NPP) hydrolysis was 0.33 mM. Its activity was inhibited non-competitively by sodium fluoride. Mercuric chloride, sodium molybdate, and copper sulfate strongly inhibited the enzyme activity. The isozyme actively hydrolyzed adenosine triphosphate and p-NPP, and partially utilized fructose-1,6-diphosphate. It did not utilize fructose-6-phosphate, glucose-1-phosphate, glucose-6-phosphate, glycerophosphate, or adenosine monophosphate.
Keywords: Acid phosphatase; Enzyme purification; Isozyme; Oryza sativa.
Introduction
Acid phosphatases (EC 3. 1. 3. 2.) occur ubiquitously among plants and animals (Hollander, 1971; Panara et al., 1990); however, their functional significance in plants is still relatively little known. Studying acid phosphatases is difficult due to their multiform occurrence in organisms, their relative nonspecificity, their small quantity, and their instability in dilute solution (Park and van Etten, 1986). Their study is also complicated by wide variations in the activity and property of isozymes between speciesand between different stages in each plant's development (Alves et al., 1994; Baes and Van Cutasem, 1993). There is also some inconsistency in nomenclature between different isozyme systems (Arnold et al., 1987; Moore et al., 1987). Only limited information is available on their specificity, localization, and physiological function (Baker and Tadakazu, 1973; Charvat and Esau, 1974; Mizuta and Suda, 1980; Chen et al., 1992).
Recent development of chromatographic methods has facilitated purification of individual isozymes of acid phosphatase. By using a combination of DEAE-Cellulose chromatography and cell fractionation, Mizuta and Suda (1980) were able to isolate twelve isozymes from bean hypocotyl. Acid phosphatase purified by a sequence of DEAE-Cellulose and/or Con A-Sepharose chromatography was also reported for tomato cultivars (Paul and Williamson, 1987), sunflower seeds (Park and Van Etten, 1986) and wheat germ (Waymack and Van Etten, 1991).
In this study, several different chromatographic methods were employed to purify one acid phosphatase
isozyme from rice seedlings, and its properties were examined.
Materials and Methods
Plant Materials
Rice (Oryza sativa Tainung 67) seeds were surface sterilized in 6% calcium hypochlorite, soaked in water for three days, and sown on plastic net moistened with nutrient solution. After germination, seedlings were transferred to the greenhouse of the Department of Horticulture, National Taiwan University. Ten-day-old seedlings were collected for enzyme extraction.
Enzyme Extraction and Purification
Rice leaves (500 g) were rapidly frozen in liquid N2, ground into powder with a mill, then soaked in 1000 ml of 50 mM buffer A (Tris-HCl, pH 7.6, 1 mM b-mercaptoethanol). The mixture was homogenized with a Polytron, filtered through four layers of cheesecloth, and centrifuged at 16,000 g for 20 min. Proteins were precipitated from the supernatant with a 40_70% saturation of ammonium sulfate, and the precipitate was dialyzed against buffer A overnight. The dialysate was loaded on to a DEAE-Sepharose Fast Flow column (Pharmacia, 26 × 700 mm) and eluted with 1000 ml of 0-1 M NaCl in a linear gradient at a flow rate of 200 ml/h. Fractions showing enzyme activity were pooled, concentrated through a YM 10 membrane (Amicon), and dialyzed against buffer B (25 mM Tris-HCl, pH=7.6, 50 ml NaCl). The dialysate was then loaded on to a Con A-Sepharose column (Bio-Rad, 12 × 40 ml). The loosely bound proteins were washed with buffer B, and the bound glycoproteins were eluted
1Corresponding author.