Bot. Bull. Acad. Sin. (1997) 38: 257-262
Mündel et al. Alternaria seedling blight of safflower, temperature/moisture
Effect of soil moisture, soil temperature and seed-borne
Alternaria carthami, on emergence of safflower
(Carthamus tinctorius L.)
H.-H. Mündel1, H.C. Huang, G.C. Kozub, and C.R.G. Daniels
Agriculture and Agri-Food Canada Research Centre, P.O. Box 3000 Main, Lethbridge, Alberta, Canada T1J 4B1
(Received May 26, 1997; Accepted July 22, 1997)
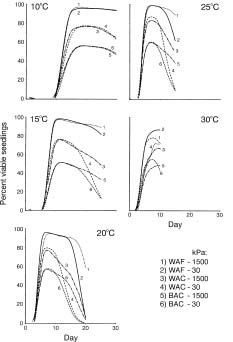
Abstract. Clean white seeds of safflower, Carthamus tinctorius L. cv. Saffire, from fields in Lethbridge, Alberta, Canada and Portage-la-Prairie, Manitoba, Canada and brown seeds infected with Alternaria carthami Chowdhury from Portage-la-Prairie were used to test the effects of soil moisture and temperature on seedling emergence. Seeds were planted in autoclaved soil, at two water potentials (30 kPa and 1500 kPa), and at five temperatures (10, 15, 20, 25, and 30°C) and rated for seedling emergence. The Alternaria carthami seeds were 32%, 82%, and 96% infected in the white seeds from Lethbridge, white seeds from Portage-la-Prairie, and brown seeds from Portage-la-Prairies, respectively. For temperatures less than 30°C, emergence of seedlings was greatest (>90%) for the white Lethbridge seed; followed by the white Portage-la-Prairie seed (75_85%); with lowest emergence (50_60%) from the brown Portage-la-Prairie seed, over both soil water potentials. While seed source had significant effects on total seedling emergence, temperature and soil water potential did not. However, post-emergence damping-off increased with higher temperature, and its incidence was greater at high soil moisture (with temperatures above 10°C).
Keywords: Alternaria carthami; Carthamus tinctorius; Seed-borne; Seedling emergence; Soil water potential; Temperature.
Introduction
Alternaria carthami Chowdhury is an important seedborne pathogen of safflower (Carthamus tinctorius L.) in western Canada (Petrie, 1974), U.S.A. (Burns, 1974), Australia (Irwin, 1976), and India (Chowdhury, 1944). Mortensen et al. (1983) found that A. carthami was pathogenic on safflower at all growth stages in Montana, causing up to 50% seed rot and seedling blight in susceptible cultivars. In Canada, Howard et al. (1989, 1990) found seedling blight in all fields of safflower surveyed in areas of southern Alberta, with incidence varying from 6 to 53% in 1988 and averaging 52% in 1989. Petrie (1974) found that A. carthami occurred in up to 95% of safflower seeds in some fields in Saskatchewan. Also seed-borne inoculum became the primary source of inoculum of leaf spot during the growing season.
Previous investigations have established that safflower seed produced in California and Arizona is generally free of seed-borne pathogens, particularly A. carthami (Burns, 1974). Seed produced in Alberta, Manitoba, Saskatchewan, and Montana, however, is often heavily contaminated with this pathogen, resulting in reduced germination and seedling vigour (Mortensen et al., 1983; Mündel, 1996). Fungicidal seed treatments have only partly reduced the incidence of seed-borne A. carthami,
with the pathogen internal to the seed coat not being killed (Irwin 1976; Mortensen et al., 1983).
Mündel et al. (1995) reported the effects of soil water potential and temperature on damping-off of safflower caused by the soil-borne pathogen Pythium sp. "group G", a form of Pythium ultimum (Huang et al., 1992). This study was undertaken under a controlled environment to determine the effects of soil moisture and temperature on damping-off and seedling blight of safflower caused by the seed-borne pathogen A. carthami.
Materials and Methods
Experiments were performed in temperature-controlled rooms to study safflower seedling emergence and survival, using cv. Saffire (Mündel et al., 1985). Seeds (achenes) of this cultivar are normally white in color. However, infection of seeds by A. carthami results in discoloration of the seed coat. Seed samples from three 1991 field variety trials were used: a) white seeds from Lethbridge, Alberta, Canada (WAF); b) hand-selected white seeds from Portage-la-Prairie, Manitoba, Canada (WAC); and c) hand-selected brown seed from the same field in Portage- la-Prairie (BAC). The germination rates of seeds from WAF, WAC, and BAC, rolled in moist paper towelling, kept at ambient room temperature, and counted after four days, were 98%, 84%, and 80%, respectively. Two soil water potentials, low stress (30 kPa) and high stress (1500 kPa), and five temperature regimes (10, 15, 20, 25, and 30°C) were applied to the experimental units.
1Corresponding author. Fax: (403) 382-3156; E-mail: muendel@em.agr.ca