Bot. Bull. Acad. Sin. (1997) 38: 267-272
Park and Chung Gene flow in Hosta
Indirect measurement of gene flow in Hosta capitata (Liliaceae)
Ki Bae Park1 and Myong Gi Chung2,3
1Department of Horticulture, Ansung National University, Ansung 456-749, The Republic of Korea
2Department of Biology, Gyeongsang National University, Chinju 660-701, The Republic of Korea
(Received March 7, 1997; Accepted July 22, 1997)
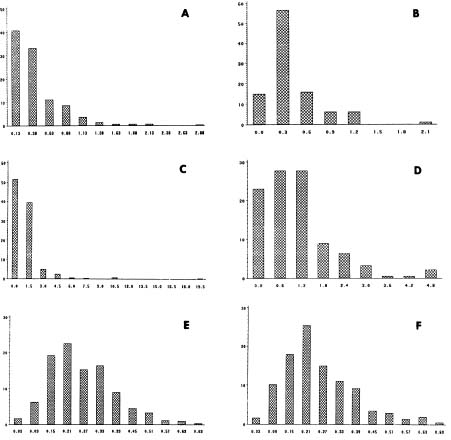
Abstract. We investigated the nearest neighbor distances between plants, pollinator flight distances, and seed dispersal distances in two natural populations of Hosta capitata to estimate indirectly the neighborhood area and size of this species. As with most insect-pollinated species, the frequencies and shapes of all three variables showed significant skewness and leptokurtosis. Means of pollinator (bumblebees and Apis) flight distances and seed dispersal distances of H. capitata were about 1 m and 25 cm, respectively. In addition, although neighborhood sizes differed (ca. 15 and 64 individuals), as did neighborhood areas (5.86 to 17.13 m2), between the two populations, these results indicated substantial spatial restriction of gene flow within them. The present results in conjunction with previous studies on fine and large scale genetic structure using allozymes as genetic markers within and among populations of H. capitata strongly indicate that gene flow is sufficiently restricted to allow stochastic effects to play an important role in determining the genetic structure of this species.
Keywords: Gene flow; Genetic drift; Hosta capitata; Pollinators; Seed dispersal.
Introduction
The effective population size (Ne) is related to the effective number of individuals in a plant population that interbreed randomly, taking into consideration population density and the variances in gene flow components. In other words, pollen travel and seed dispersal are two components of gene flow in plant populations. These patterns in conjunction with the varying density of individuals determine the Ne. Neighborhood is the area within a plant population in which panmixis, that is, random gene exchange via pollen and seed dispersal occurs (Wright, 1946). Levels of gene flow affect Ne and the magnitude of Ne determines the magnitude and direction of the evolutionary factors acting on the genetic variation within populations (Wright, 1938). For example, high rates of gene flow over large geographical areas increases Ne and can overshadow the influence of natural selection or genetic drift, reducing genetic differentiation. Thus, accurate measures of gene flow have been of continued interest to evolutionary biologists. However, relatively few studies of neighborhood sizes in plants have been reported in the plant population biology literature (Richards, 1986), partly due to two practical reasons or problems, as suggested by Richards (1986): 1) measurements of gene movement tend to vary considerably spatially and temporally and 2) estimations of Ne are largely dependent on plant density.
Hosta capitata (Koidz.) Nakai (Liliaceae), an herbaceous perennial, occurs in the southern Korean Peninsula and southwestern Japan (Fujita, 1976; Chung and Kim,
1991). In Korea, most populations of the species are relatively small and isolated compared with other Korean hostas (Chung et al., 1991) and are found in pine-oak forest understories and in grasslands in mountainous regions. As the specific epithet indicates, 3_12 flowers are clustered on the top of a scape. Flowers are visited by bees (Bombus diversus diversus and Apis mellifera; Chung, 1994a). Although studies on the breeding system are not available, H. capitata appears to be predominantly outcrossing because most of the flowers have pronounced spatial separation (herkogamy) of mature anthers and stigmas impeding self-pollination. The fruit of this species is a cylindrical capsule, with 10_30 small (3.0_5.5 mm) seeds in each capsule. The seeds are winged. Previous studies have reported that populations of H. capitata maintain low levels of allozyme diversity and there is high allozyme divergence among populations (Chung et al., 1991; Chung, 1994a). In an effort to understand factors affecting levels of genetic variation within populations and the population genetic structure of H. capitata, the senior author has investigated spatial genetic structure within and among its populations in Korea using spatial autocorrelation analysis (Chung, 1996, 1997). These studies indicate that gene flow within a circumscribed area and among populations is considerably low, suggesting that neighborhood size and area are small. As the physical distances of dispersal should be related to the density of individuals in order to relate to the physical scale of spatial structure (Epperson, 1995), information (e.g., Wright's neighborhood size and area) concerning the standardized dispersal in populations of H. capitata is necessary. In this study, nearest neighbor distances between plants of the insect-pollinated, H. capitata were measured. In addition, pollinator flight and seed dispersal distances were also measured in two natural popu
3Corresponding author. Fax: +82-591-54-0086.