Bot. Bull. Acad. Sin. (1997) 38: 277-284
Morris and Lammers Pollen of Codonopsis
Circumscription of Codonopsis and the allied genera Campanumoea and Leptocodon (Campanulaceae:
Campanuloideae). I. Palynological data
Kari E. Morris1,2 and Thomas G. Lammers2,3
1Department of Biological Sciences, University of Illinois at Chicago, Chicago, IL 60608, USA
2Department of Botany, Center for Evolutionary and Environmental Biology, Field Museum of Natural History, Chicago, IL 60605-2496, USA
(Received January 28, 1997; Accepted May 16, 1997)
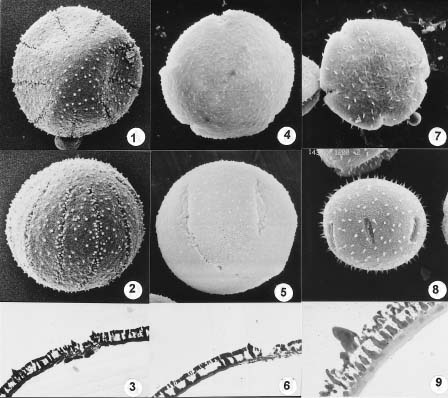
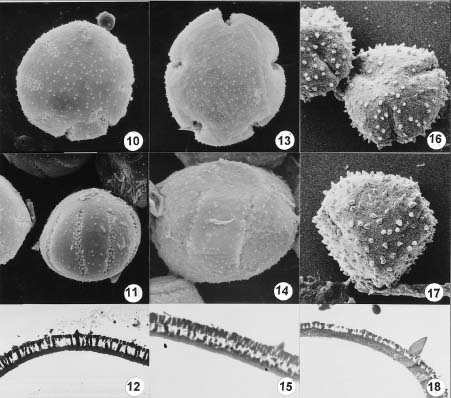
Abstract. Pollen from 23 accessions of Codonopsis and the allied genera Campanumoea and Leptocodon was examined via light microscopy, scanning electron microscopy, and transmission electron microscopy. All grains examined were of four types. Type I grains (found in all accessions of Campanumoea sect. Campanumoea, Codonopsis subg. Codonopsis, and Leptocodon) were oblate sphaeroidal, circular in polar outline, 4_10-colpate, with long colpi, short spinules, and thin nexine. Type II grains (found in all accessions of Campanumoea sect. Cyclocodon) were oblate sphaeroidal, semi-angular in polar outline, 3-colporate, with long colpi, long spinules, and thick nexine. Type III grains (found in all accessions of Codonopsis subg. Pseudocodonopsis) resembled Type I grains, but had the long spinules characteristic of Type II; further, the colpi were much shorter than in Type I and II. Type IV grains (found in all accessions of Codonopsis subg. Obconicicapsula) likewise resembled Type I grains, but had colpi of a length intermediate between Type I and Type III. The uniformity of pollen morphology in Campanumoea sect. Campanumoea, Codonopsis subgen. Codonopsis, and Leptocodon might be interpreted as supporting their merger as a single genus, while the distinctness of Type I and Type II pollen might be interpreted as supporting the recognition of Cyclocodon as a genus distinct from Codonopsis. However, the palynological data are less clear as to the disposition of Codonopsis subg. Pseudocodonopsis and subg. Obconicicapsula, and a realignment of generic boundaries must await completion of studies involving additional data sets.
Keywords: Campanulaceae; Campanuloideae; Campanumoea; Codonopsis; Cyclocodon; Electron microscopy; Leptocodon; Pollen; Palynology; Ultrastructure.
Introduction
Codonopsis* (Campanulaceae: Campanuloideae) comprises 55 species endemic to eastern Asia, from Kamchatka and Japan south to Afghanistan, Pakistan, the Himalayas, southern China, and Taiwan (Fedorov, 1957; Rechinger and Schiman-Czeika, 1965; Hong, 1983; Shimizu, 1993). The plants are perennial with tuberous roots; scandent, twining, or less commonly erect or ascending herbaceous stems; solitary or paired axillary or terminal flowers; 3_5-locular ovary with a 3_5-lobed stigma; and capsular fruits loculicidally dehiscent at apex; most have a pungent skunk-like or `foxy' odor of uncertain chemistry. Chromosome number has been determined for eight species; all show n = 8 (Rosén, 1931; Gadella, 1966; Lee, 1967; Zhukova, 1967; Krasnoborov et al., 1980; Nishikawa, 1985; Sui et al., 1985; Jee et al., 1989; Yoo and Lee, 1989; Ge and Wang, 1990). The most recent classification of the genus (Hong, 1983) recognized three subgenera: Codonopsis, Obconicicapsula, and Pseudocodonopsis.
Some authors (Moeliono and Tuyn, 1960; Van Thuan, 1969; Lammers, 1992) have expanded the circumscription of Codonopsis to include Campanumoea. This genus of five species is likewise endemic to eastern Asia, though its distribution extends farther south than Codonopsis, to tropical India, Indochina, the Philippines, Malaysia, Indonesia, and New Guinea (Moeliono and Tuyn, 1960). Campanumoea indeed resembles Codonopsis, but differs in its baccate fruit; furthermore, it lacks the distinctive foxy odor found in most Codonopsis, and the ovary is often 6-locular with a 6-lobed stigma. The species have been divided (Clarke, 1881; Tsoong, 1935; Hong, 1983; Shimizu, 1993) between two sections: Campanumoea and Cyclocodon. According to Hong (1995), the chromosome number of the former is n = 8, while that of the latter is n = 9.
Recently, Grey-Wilson (1990) has proposed that Codonopsis be enlarged further through the incorporation of Leptocodon, a genus of two species endemic to the Himalayas, from India to Burma. Leptocodon resembles Codonopsis overall, but differs in the possession of a unique set of five glands atop the ovary, alternate with the stamens. The chromosome number of Leptocodon is unknown.
3Corresponding author. E-mail: lammers@fmppr.fmnh.org
*Authorities for all scientific names will be found in Table 1.