Bot. Bull. Acad. Sin. (1998) 39: 9_15
Kung et al. A cold-induced gene from pea
Characterization of a pea gene responsive to low temperature
Cheng-Che S. Kung, Kai-Wun Yeh, Chu-Yung Lin, and Yih-Ming Chen1
Department of Botany, National Taiwan University, Taipei, Taiwan 106, Republic of China
(Received April 12, 1997; Accepted September 11, 1997)
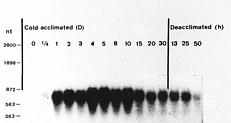
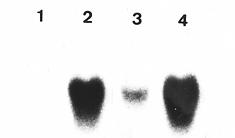
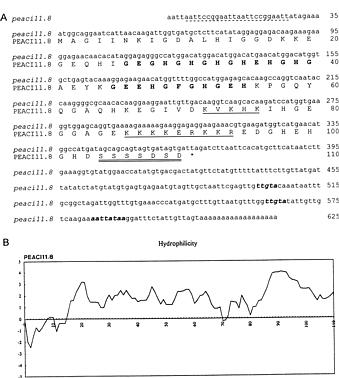
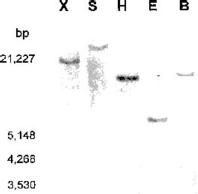
Abstract. A cold-induced gene, peaci11.8, was isolated by differential screening of a subtracted cDNA library from cold-acclimated etiolated seedlings of pea (Pisum sativum L.). The expression of peaci11.8 gene was investigated. The accumulation of the transcripts began within 6 h of cold treatment and peaked between 4 and 10 days of treatment. The expression of this gene was rapidly down-regulated after the acclimated seedlings were transferred to ambient temperature. This indicated the correlation of the peaci11.8 gene expression and cold treatment. Further studies showed that the expression was also highly stimulated by water stress. The amino acid sequence deduced from the nucleotide sequence of peaci11.8 shared significant homology with several plant cold-induced genes. In addition, the sequence analysis revealed the presence of two common features of many late embryogenesis abundant (LEA) proteins, the lysine-rich motif and the serine cluster. The availibility of peaci11.8 will facilitate the structure-function studies of the cold-inducible gene product.
Keywords: Cold acclimation; Gene expression; Pisum sativum.
Abbreviations: LEA, late embryogenesis abundant; NTS, nuclear-targeting signal; ORF, open reading frame; SSC, 150 mM NaCl/15 mM sodium citrate.
Introduction
Temperature is one of the major environmental factors affecting nutrient distribution, growth, and development of plants. Cold-sensitive plants, especially those of tropical or subtropical origins, are readily injured or killed at low, nonfreezing temperatures (Levitt, 1980). In contrast, plants of temperate areas are more tolerant to freezing when previously exposed to low positive temperatures, a process known as cold acclimation. Comparative analyses have revealed that various biochemical and physiological changes occur during cold acclimation, including alterations in lipid and carbohydrate compositions as well as protein contents and enzyme activities (Guy, 1990; Thomashow, 1990).
It has been well-demonstrated that changes in gene expression and protein synthesis are associated with induction of freezing tolerance during cold acclimation (Gilmour et al., 1988; Guy et al., 1985). Thus far, cold-acclimation-specific transcripts have been identified in various plant species including Arabidopsis thaliana (Kurkela and Franck, 1990), Brassica napus (Orr et al., 1992), Hordeum vulgare (Cativelli and Bartels, 1990), Medicago sativa (Wolfraim and Dhindsa, 1993), Oryza sativa (Aguan et al., 1991), Spinacia oleracea (Neven et al., 1993), and Triticum aestivum (Chauvin et al., 1993). Although functions of these proteins and the molecular mechanisms regulating expression of these genes remain largely unknown, they are thought to be closely related to cold acclimation.
Recently, we found a 19-kDa thylakoid membrane-associating protein increased remarkably when pea (Pisum sativum) plants were exposed to low temperature (Ma et al., 1990). To obtain a better insight into the mechanisms of cellular acclimation, efforts were made to isolate and characterize cDNA clones corresponding to cold-induced transcripts from pea. In this report, we describe the cloning and characterization of a novel gene of pea that responds to low temperature.
Materials and Methods
Plant Material and Treatments
Seeds of pea (Pisum sativum L. cv. Taichung No. 9) were rinsed with tap water and germinated in rolls of moist paper towel at ambient temperature (28°C) in a dark growth chamber. Two-day-old seedlings were either harvested without cotyledons or used for cold (4°C) treatment. For cold acclimation, seedlings were rinsed with ice-cold water, then transferred to cold in dark and harvested at prescribed time intervals. For deacclimation, seedlings acclimated for 4 days were transferred back to ambient temperature and harvested at a specified time. For other stresses, seedlings were harvested 1 day after being transferred to 20% PEG 6,000 or 100 mM ABA at ambient temperature, respectively.
RNA Isolation
Total and poly(A)+ RNA were extracted according to the method of Zurfluh and Guilfoyle (1982), except that the poly(A)+ RNA was precipitated in the presence of 0.3 M ammonium acetate and two volumes of ethanol.
1Corresponding author. Fax: 886-2-23919096; E-mail: chuyung@ccms.ntu.edu.tw