Bot. Bull. Acad. Sin. (1998) 39: 17_21
Yang et al. Genetic polymorphism in Capsella bursa-pastoris
Genetic polymorphism of seven populations of Capsella bursa-pastoris based on RAPD markers
Yau-Wen Yang1,3, Warren H.J. Kuo2, and Teck Hiang Wong1
1Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
2Department of Agronomy, National Taiwan University, Taiwan 106, Republic of China
(Received June 16, 1997; Accepted October 22, 1997)
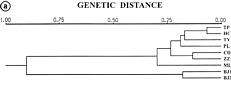
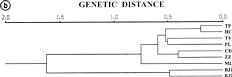
Abstract. Capsella bursa-pastoris is considered to be a potential vegetable crop. Together with two accessions of mustard (Brassica juncea) as outgroups, seven populations of C. bursa-postoris were used to study genetic polymorphism using random amplified polymorphic DNA (RAPD) markers. Using 12 primers, a total of 158 RAPD markers were selected among these populations. Among them, 91 are either specific for C. bursa-postris or present in both C. bursa-postoris and B. juncea. In Capsella populations, 40 RAPD markers are polymorphic, 5 markers are population specific, and 46 markers are monomorphic with variation of banding density. Each RAPD marker was treated as either having unweighted character or weighted character. Using UPGMA for cluster analysis, the polymorphic tree structures based on unweighted and weighted character have a similar phylogenetic relationship among these tested populations. With one exception, the phylogenetic relationship of Taiwan origin is correlated with the geographical distribution of these populations. These results may provide us information of genetic variation to be used in a varietal development program and to find specific markers linked to important traits.
Keywords: Capsella bursa-pastrois; Genetic polymorphism; RAPDs.
Introduction
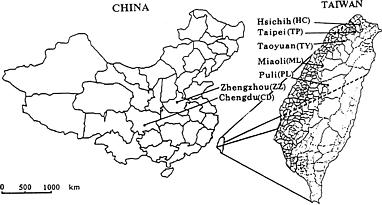
Capsella bursa-pastoris is a wild grass and the only species of the genus Capsella in Taiwan (Yang, 1996). With its pleasant, distinctive flavor, it has potential for cultivation as a vegetable crop (Kuo and Chen, 1995). Due to the small leaf size and early flowering of the existing Taiwan populations, high yields and economical production on a large scale are difficult. Two populations from the middle of China have been introduced into Taiwan. With their large leaves and late flowering habit these populations are good candidates for generating new varieties, or F1 hybrids, with Taiwan populations. Information on genetic variabilities and relationships among these different populations may aid in an ecotype identification and future varietal development program.
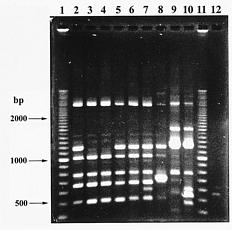
With the advent of molecular biology, random amplified polymorphic DNA (RAPD) markers generated by polymerase chain reaction (PCR) with single 10base oligonucleotide primers of arbitrary sequence have proved to be an easier, quicker way for detecting polymorphism at the DNA level compared with restriction fragment length polymorphism (RFLP) (Welsh and McClelland, 1990; Williams et al., 1990). However, the principal disadvantage of RAPDs is that they are usually dominant markers, and reproducibility of DNA banding patterns can be affected by different concentrations of reaction components and cycle conditions (dos Santos et al., 1994). In
Brassica and its related genera, RAPD markers have been used successfully for identification and phylogenetic relationship among and within species (Demeke et al., 1992; Kresovich et al., 1992; Hu and Quiros, 1991; Ren et al., 1995; Quiros et al., 1991; dos Santos et al., 1994; Thormann et al., 1994). F1 plants of B. juncea show a positive relationship between yield and genetic distance (Jain et al., 1994). Some RAPD markers can even be linked to special traits, like the fertility restorer gene for use against cytoplasmic male sterility in the F1 seed production of rapeseed (Delourme et al., 1994).
This study has two objectives. The first is to evaluate if there is genetic variation among collected populations using RAPD markers since no work has been done on these plants yet. The second is to see how different evaluation methods of RAPD banding patterns in genetic distance can affect the phylogenetic relationship among these populations. Usually, RAPD bands were treated as either "present" or "absent" without considering banding density. This study was done to see whether assigning scores to different band densities affects the analytical result. Here, we adopted two methods for estimating genetic distance among the tested populations. In the first, each DNA band was treated as a unit character, and each population was scored for the presence or absence of band, respectively. Genetic Similarity (GS) was analyzed using the equation (Jaccard, 1908): GS = (Nab)/(Na+Nb-Nab), where Nab is the number of shared fragments between populations a and b, Na is the number of scored fragments of population a, and Nb the number of scored fragments of population b.
3Corresponding author.