Ann et al. Mating type and pathogenicity of Phytophthora infestans in Taiwan
Bot. Bull. Acad. Sin. (1998) 39: 33_37
Mating type distribution and pathogenicity of Phytophthora infestans in Taiwan
P. J. Ann1, T. T. Chang2,3 and L. L. Chern2
1Chia-Yi Agricultural Experiment Station, Chia-yi, Taiwan, Republic of China
2Division of Forest Protection, Taiwan Forestry Research Institute, 53 Nan-Hai Road, Taipei, Taiwan, Republic of China
(Received April 22, 1997; Accepted November 18, 1997)
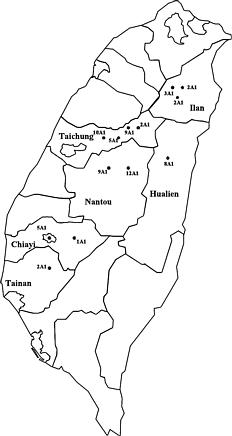
Abstract. Taiwanese isolates of Phytophthora infestans were examined for mating types and pathogenicity to tomato and potato. A total of 70 isolates, including 68 isolates from tomato and 2 isolates from potato, from six counties of Taiwan were all A1 mating type. Three tomato isolates of P. infestans caused only leaf blight of tomato, while two isolates obtained from potato were able to cause the disease both on potato and tomato, indicating that host-specific strains of P. infestans occur in Taiwan.
Keywords: Host specificity; Mating type; Phytophthora infestans; Potato; Taiwan; Tomato.
Introduction
The fungus Phytophthora infestans (Montagne) de Bary is a heterothallic oomycete with two compatibility mating types designated as A1 and A2 (Gallegly and Galindo, 1960). This species has been known to exist in Taiwan since the early 1900s, causing late blight of potato and tomato (Kawakami and Suzuki, 1908; Sawada, 1919). Three tomato isolates of the fungus belonging to the A1 mating type from Taiwan have been analyzed for genetic variation at two allozyme loci, glucose-6-phosphate isomerase and peptidase, and for nuclear DNA haplotype. The three isolates all had a genotype identifical with the one previously found throughout the world, designated as US-1 (Koh et al., 1994). However, the mating type composition and pathogenicity to tomato and potato within the P. infestans population of Taiwan have not previously been documented. Results of an investigation into these aspects are reported in this paper.
Materials and Methods
Isolation of Phytophthora infestans
Samples of blighted tomato and potato leaves were collected from tomato and potato fields in 1993_1996. The blighted leaves were kept in a plastic bag for 2_4 days at 20°C to promote sporangia formation. The sporangia were picked off the blighted leaves using a sterile needle and spread on rye B agar (Frinking et al., 1987) or green pea agar (Shattock et al., 1990) amended with 100 ppm ampicillin, 50 ppm mycostatin, and 10 ppm
pentachloronitrobenzene after autoclaving. After incubation at 20°C for 5_10 days, mycelia of P. infestans germinating from sporangia were transferred to rye B agar or green pea agar.
Characteristics of Phytophthora infestans
Sporangia for measurement were obtained from cultures of P. infestans grown on rye B agar for 2_3 weeks in darkness. P. infestans isolates were grown on rye B agar for 5_10 days. Agar discs (5 mm in diameter) cut from the periphery of the colonies with a sterile cork borer were used to inoculate plates. Each rye B agar plate was inoculated with one disc of inoculum placed at the margin of the plate. Three tomato isolates and one potato isolate were used for characteristics. To determine the effect of temperature on the growth of each isolate, inoculated agar plates were incubated at 10, 12, 15, 18, 20, 24, 25, 26 and 27°C in darkness. The linear growth of mycelium was measured 10 days after inoculation. Four plates were used for each temperature, and the experiment was performed twice.
Determination of Mating Types
The mating type of each culture of P. infestans was determined by pairing a small piece (ca. 3 × 3 × 3 mm) of a culture to be tested with the same size of the A1 tester culture (isolate 533) or the A2 tester (isolate IB905) on a rye A agar block (ca. 15 × 10 × 3 mm) in a petri dish. Ten blocks were placed in a petri dish at equal distance along the edge. After incubation at 20°C in darkness for 10_14 days, agar blocks were examined microscopically. The isolates forming oospores when paired with the A2 tester were designated as A1. Similarly, isolates forming oospores when paired with A1 tester were designated A2.
3Corresponding author. Tel & Fax:+886-2-23891419; E-mail: ttchang@serv.tfri.gov.tw