Bot. Bull. Acad. Sin. (1998) 39: 39_45
Kao and Chang d13C and nutrient contents of Kandelia candel
Stable carbon isotope ratio and nutrient contents of the Kandelia candel mangrove populations of different growth forms
Wen-Yuan Kao1 and Kuo-Wei Chang
Institute of Botany, Academia Sinica, Taipei, Taiwan, Republic of China
(Received September 17, 1997; Accepted November 26, 1997)
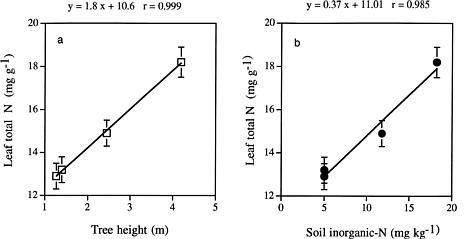
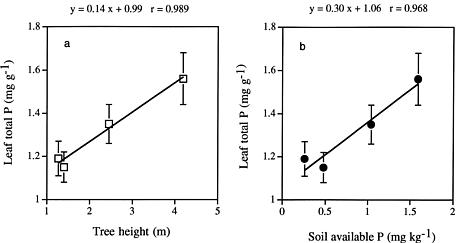
Abstract. Kandelia candel (L.) Druce, the dominant mangrove species on the west coast of northern Taiwan, has three distinct growth forms. To elucidate whether the growth of the dwarf form of K. candel is related to nutrient and water availability, we analyzed nutrient contents (N, P, K, Ca, Mg, Na) and carbon isotope ratios (d13C) of leaves of four K. candel populations differing in canopy height. The objectives of the study were to investigate whether the dwarf K. candel populations have higher long-term water use efficiency and whether nutrient limitation is related to the growth of the dwarf form of mangrove plants. Leaves of dwarf mangrove populations had significantly higher d13C values (1_1.5 higher) and thus higher long-term integrated water use efficiency than the taller mangrove populations. N and P contents were significantly lower in leaves and soils of the dwarf mangrove populations than in those of medium height and tall mangrove populations. In contrast, no significant difference was found in K, Na, Ca and Mg contents among leaves of the four mangrove populations. It is proposed that water and nutrient availability, especially of nitrogen and phosphorus, limit the growth of K. candel.
Keywords: Kandelia candel; Mangrove; Nitrogen content; Phosphorus content; Stable carbon isotope ratio; Water use efficiency.
Introduction
Mangrove are a group of trees or shrubs that grow in the tidal waters of tropical and subtropical coastlines. Though mangrove are able to colonize saline habitats, factors such as high salinity, poor aeration (Davis, 1940), waterlogging (Egler, 1952), compacted peat (Craighead, 1971), and nutrient limitations (Feller, 1995; Lugo and Snedaker, 1974) have been thought to limit their growth. In addition, Lin and Sternberg (Lin and Sternberg, 1992a, b), using stable isotope techniques to compare the ecophysiology of scrub and fringe mangroves in Florida, USA, concluded that water availability and fluctuations in salinity could also limit the growth of mangrove plants.
Most of the above studies were conducted on Rhizophora mangle (Feller, 1995; Lin and Sternberg, 1992a,b; Lugo and Snedaker, 1974), Avicennia spp. (Lin and Sternberg, 1992a; Popp, 1984), Aegialitis annulata (Popp, 1984), and Languncularia spp. (Lin and Sternberg, 1992a). Kandelia candel is the dominant mangrove species in the west coast of northern Taiwan (Liu, 1982). Kandelia candel in the Chu-wei area of northern Taiwan exhibits three distinct growth forms: The average tree height of dwarf populations is about 1 m; the tall population is 4 m or greater; and the medium height population is 2 to 3 m. Factors responsible for the different growth
forms of this mangrove species have not been studied. Huang (1983) found significant differences in soil nutrient content and salinity at six locations within the Chu-wei area. We also observed that dwarf form populations are located at higher elevations where there is less flooding. These observations suggest that the dwarf form of K. candel might be related to nutrient and/or water availability. In the present study, we focused on these two factors.
We thus conducted a survey to compare the long-term water use efficiency (WUE) and nutrient characteristics of leaves of K. candel plants from populations of different growth forms. Because there is a negative linear relationship between leaf stable carbon isotope ratio and the ratio of intercellular to ambient CO2 concentration (Ci/Ca) (Farquhar et al., 1982), stable carbon isotope ratios are often used to indicate the long-term integrated WUE of plants (Farquhar et al., 1989). Hence, we used the technique analyzing leaf carbon isotope ratio to determine the long-term WUE of the mangrove plants in these populations. In addition, we compared nutrients, N, P, K, Ca, Na, and Mg of leaves of the mangrove populations with different growth forms. The concentrations of inorganic nitrogen (ammonia-N and nitrate-N) and available phosphorus of the corresponding soil samples were also analyzed. There are two specific objectives of this study: (1) to investigate whether dwarf form Kandelia candel populations have higher long-term water use efficiency; and (2) to evaluate whether nutrient limitations can explain variations in size among mangrove populations in the study site.
1Corresponding author. Fax: 886-2-782-7954; E-mail: bowykao@ccvax.sinica.edu.tw