Bot. Bull. Acad. Sin. (1998) 39: 57_68
Chiang Andreaea of Taiwan
Taxonomic revision of Andreaea (Mosses, Andreaeaceae) of Taiwan
Tzen-Yuh Chiang
Department of Biology, Cheng-Kung University, Tainan, Taiwan 700, Republic of China
(Received June 17, 1997; Accepted August 6, 1997)
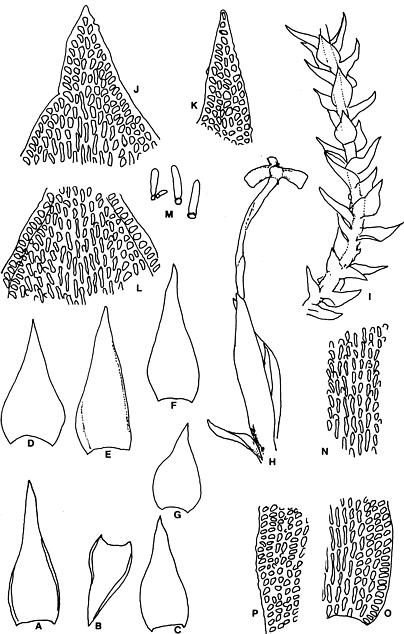
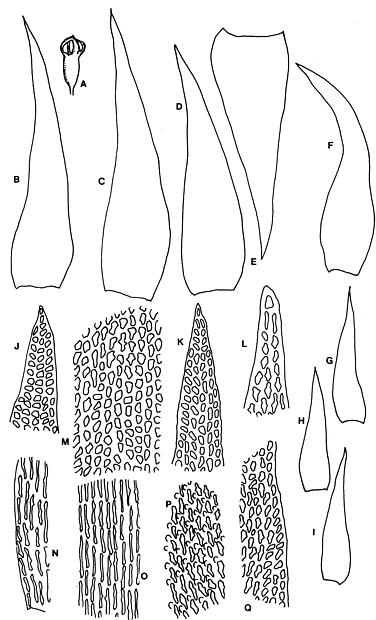
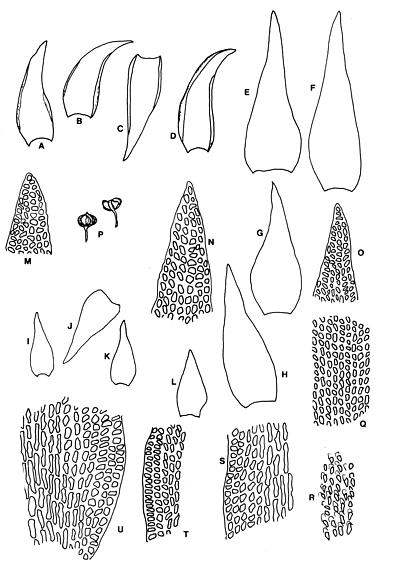
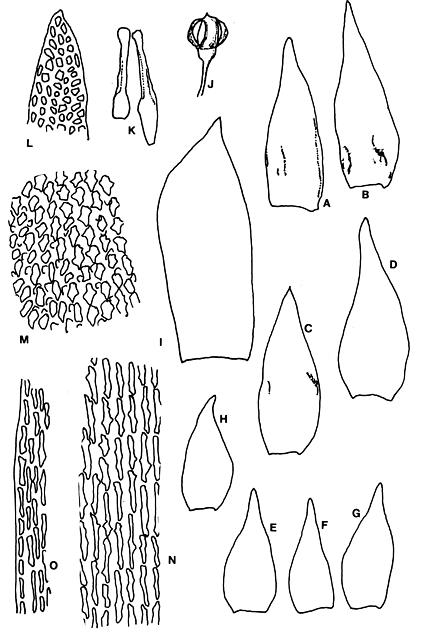
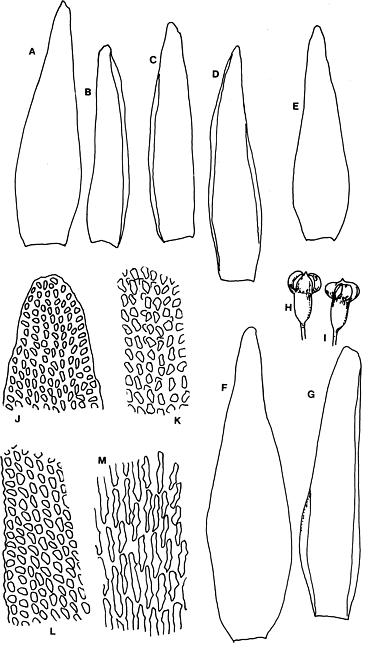
Abstract. Six taxa of Andreaea of Taiwan are revised taxonomically with one new species, A. taiwanensis sp. nov., and two new additions, i.e., A. wangiana Chen and A. rupestris Hedw. A. commutata C. Muell. is synonymized to A. rupestris var. fauriei (Besch.) Tak. Four species have capsules that split from base to tip: A. taiwanensis, A. mutabilis, A. rupestris var. rupestris, and A. rupestris var. fauriei. A. taiwanensis, closely related to A. mutabilis, is distinguishable from other Andreaeae by its large, loose leaves and rectangular to linear laminal cells. A. mutabilis is distinguished from the relatives A. rupestris var. rupestris and var. fauriei by the isodiametric marginal, basal cells. A. morrisonensis and A. wangiana belong to another group that has capsules with dehiscence in the upper half. A distinctively obtuse leaf apex distinguishes A. wangiana from A. morrisonensis.
Keywords: Andreaea; A. taiwanensis sp. nov.; Dehiscence of capsule; Taxonomy; Taiwan.
Introduction
Andreaeaceae, a monotypic family with genus Andreaea, are the typical cool-temperate and subpolar mosses, with distribution extending to the alpine tundra of tropics or subtropics (Murray, 1988). In East Asia, As2 region of van der Wijk et al. (1967), five species and one subspecies have been recognized after a series of taxonomic revisions (Chen and Wan, 1958; Noguchi, 1987; Cao and Gao, 1995; also cf. van der Wijk et al., 1969; Crosby et al., 1992). A trend of the reduction of endemic species has become noticeable when most taxa were better understood. For examples, A. likiangensis Chen and A. mamillosula Chen, previous Chinese endemic species (Chen and Wan, 1958), were lately synonymized into A. rupestris Hedw. and A. rupestris var. fauriei (Besch.) Tak. respectively; A. yunnanensis Broth. (=Didymodon nigrescens Mitt.; cf. Schultze-Motel, 1970; Chiang and Kuo, 1989) and A. kashyapii Dix. [=Didymodon subandreaeoides (Kindb.) Zander] were excluded from Andreaea (Cao and Gao, 1995). Low ratio of endemism in Andreaea, such as only 7.7% in flora of Britain and Ireland (Murray, 1988), is believed to be ascribed to the long-distance dispersal of spores, which homogeneonizes the genetic variation between populations (cf. Chiang, 1997).
Three species of Andreaea were previously reported from Taiwan (cf. Chuang, 1973; Lin, 1988). Horikawa (1934) recorded the first taxon, A. fauriei Besch. (=A. rupestris var. fauriei), to this island. A. morrisonensis Noguchi, an endemic species, was found on the summit of the highest mountain, Mt. Yushan (Noguchi, 1936). Chuang (1973) published another new species, A.
hohuanensis, which was later synonymized to A. mutabilis Hook. f. & Wils. (Murray, 1988). Two more species, i.e., A. rupestris Hedw. and A. sinuosa B. M. Murray, were mentioned on a list of the flora of Mt. Yushan (Chiang, 1989), however, without citing any voucher specimens.
Interestingly, only taxa of the section Andreaea (subgenus Andreaea), ecostate species, occur in Taiwan and mainland China. Although A. rothii Web. & Mohr., a costate species of section Nerviae Card. ex. Broth. (subgenus Andreaea), appeared on Redfearn and Wu's (1986) Catalog of the Mosses of China, it has lately been excluded from the flora of China (Cao and Gao, 1995). In Asia section Nerviae with A. rigida Wils. (Gangulee, 1969) and A. subulata Harv. (Eddy, 1988) is found in India and Malesia, respectively; and another costate taxon, section Chasmocalyx Lindb. ex Braithw. (subgenus Chasmocalyx) with A. nivalis Hook., occurs in Japan only.
Species of Andreaea are locally abundant and dominant in the alpine tundra of Taiwan (Chiang, 1989), unlike the "infrequent occurrence of species in mainland China" stated by Cao and Gao (1995). Andreaea seemed rare on this island according to the previous records. For example, only one specimen of each species was examined in Chuang's (1973) revision. However this rarity proved illusive when the flora became better understood. The poor samplings were mostly caused by the difficulty of access to the high mountains as well as the indifference of collectors to the tiny mosses. Most Andreaeae grow on arid habitats and show a high polymorphism of morphological traits. The aims of this study are to revise the Andreaea based on the materials collected from high mountains of this island and to compare them with the taxa of the neighboring areas. Six taxa are studied with one new species, A. taiwanensis Chiang and two new additions, i.e., A. rupestris and A. wangiana Chen, to Taiwan.
1Corresponding author. Fax: 011-886-6-2742583; E-mail: tychiang@mail.ncku.edu.tw