Hou et al. Chitinase of sweet potato
Bot. Bull. Acad. Sin. (1998) 39: 93-97
Chitinase activity of sweet potato (Ipomoea batatas [L.] Lam var. Tainong 57)
Wen-Chi Hou1, Ying-Chou Chen2 and Yaw-Huei Lin1,3
1Institute of Botany, Academia Sinica, Nankang, Taipei 115, Taiwan, Republic of China
2Department of Bioengineering, Tatung Institute of Technology, Taipei 115, Taiwan, Republic of China
(Received September 19, 1997; Accepted February 12, 1998)
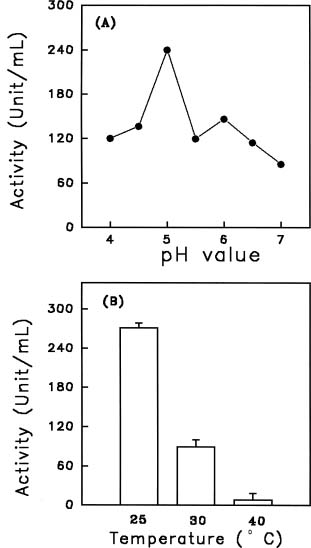
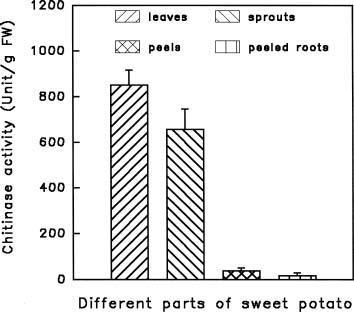
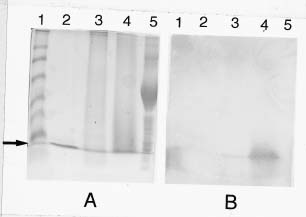
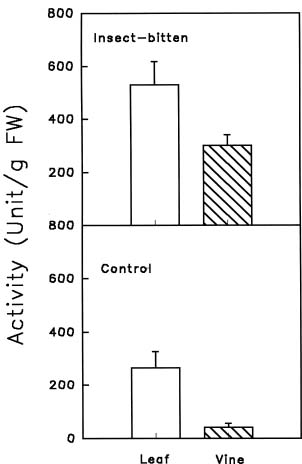
Abstract. Chitinase activity was detected in the crude extracts of different parts of sweet potato (Ipomoea batatas [L.] Lam var. Tainong 57) using commercial deacetyl glycol chitin as a substrate. The activity of chitinase in crude extracts, which was expressed as units/g fresh weight, was arranged in decending order of leaves > sprouts > peels of storage roots > peeled storage roots. Under field conditions, the chitinase activity of leaves was higher than vines. The chitinase activity of insect-bitten leaves was higher than that of the controls, and the chitinase activity of vines attached to insect-bitten leaves was also higher than that of vines attached to intact leaves. The molecular weight of main chitinase determined by activity staining was about 16 kDa. The optimum pH was 5.0 and the optimum temperature of chitinase activity was 25°C.
Keywords: Chitinase; Defense against pests; Ipomoea batatas.
Abbreviations: CBB R-250, coomassie brilliant blue R-250; GlcNAc, N-acetylglucosamine; PAGE, polyacrylamide gel electrophoresis; PVP, polyvinyl polypyrrolidone; SDS, sodium dodecyl sulfate; TCA, trichloroacetic acid
Introduction
Chitinases (EC 3.2.1.14) catalyze the hydrolysis of chitin, a linear homopolymer of b-1,4-linked N-acetylglucosamine (GlcNAc) residues. No substrates for this group of enzyme have been identified in plants, whereas chitin is commonly a component of fungal cell walls and the exoskeleton of arthropods, organisms which include many important pathogens and pests. Plants respond to attack by microorganisms, insects, and animals by the induction of sets of genes encoding diverse proteins, many of which are believed to play a role in defense (Bol et al., 1990; Bowles, 1990; Collinge et al.,1993; Dixon and Harrison, 1990; Linthorst, 1991; Punja and Zhang, 1993). The most marvelous of these proteins is chitinase. It has been demonstrated that enhanced chitinase in transgenic plants can indeed reduce the damage caused by pathogens (Broglie et al., 1991).
Most plant chitinases belong to endochitinases that randomly hydrolyze chitin polymers. They have been purified from barely (Leah et al., 1991; Swegle et al., 1989), bean (Abeles et al., 1970; Boller et al., 1983; Broglie et al., 1986), cabbage (Chang et al., 1992), corn (Lin et al., 1982), pea (Vad et al., 1991), potato (Gaynor, 1988), rice (Huang et al., 1991; Zhu and Lamb, 1991), rye (Yamaggami and Funatsu, 1993), soybean (Wadsworth and Zikakis, 1984), tobacco (Shinshi et al., 1990), wheat (Molando et al., 1979), and yam (Tsuyoshi et al., 1984). Plant chitinases fall into three classes based on primary structures (Shinshi et al., 1990). Some plant chitinases
also show lysozyme activity, which hydrolyzes the b-1,4-linkages between N-acetylmuramic acid and GlcNAc residues in peptidoglucan (Roberts and Selitrennikoff, 1988). In this preliminary report, we detected chitinase activities in different parts of sweet potato (Ipomoea batatas [L.] Lam var. Tainong 57), including leaves, sprouts, vines, peels and peeled storage roots. These results will be helpful for further investigations of sweet potato chitinases.
Materials and Methods
Materials
Fresh storage roots of sweet potato (Ipomoea batatas [L.] Lam var. Tainong 57) were purchased from a local market. After being cleaned with water, the roots were placed in a thermostated (30°C ) growth chamber in the dark and sprayed with water twice a day. Etiolated sprouts of 3_5 cm length were collected for further use. Leaves and vines of the same variety were kindly provided by Dr. C. H. Cheng (Provincial Agricultural Research Institute, Chia-Yi branch). Cut vines of sweet potato variety Tainong 57 were planted with five other varieties in Sep. 1996 in a randomized block design (4 repeat blocks, 10 m2/block). Number 39 complex fertilizer (N : P2O5 : K2O = 12 : 18 : 12) of the Taiwan Fertilizer Company was applied (0.28 kg per block) both basally and two months after planting. Both intact and insect-bitten leaves with attached vines were collected (May 19, 1997). Leaves and vines were weighed immediately and put in an envelope and frozen in liquid nitrogen in situ. The frozen samples were kept at -70°C for further use.
3Corresponding author. Fax: +886-2-2782-7954.