Botha et al. Russian wheat aphid infestation, exogenous ethylene, mechanical wounding, and chitinase isoenzymes
Bot. Bull. Acad. Sin. (1998) 39: 99_106
Chitinase isoenzymes in near-isogenic wheat lines challenged with Russian wheat aphid, exogenous ethylene, and mechanical wounding
A-M. Botha1,3,4, M.A.C. Nagel1, A.J. Van der Westhuizen1, and F.C. Botha2
1Department of Botany and Genetics, Faculty of Science, University of the Orange Free State, P. O. Box 339, Bloemfontein, 9300, South Africa
2Department of Biotechnology, SASEX, Mount Edgecombe, 4300, South Africa
(Received May 19, 1997; Accepted December 3, 1997)
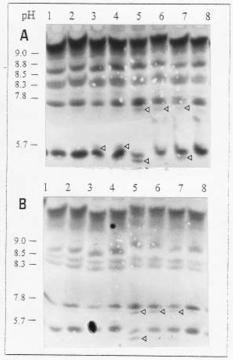
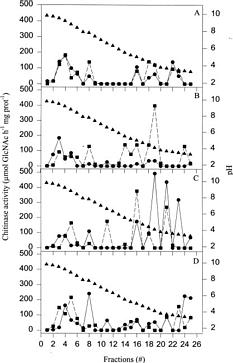
Abstract. Russian wheat aphid (RWA) infestation, ethylene exposure, and mechanical wounding resulted in differential expression of chitinase (EC 3.2.1.14) isoenzymes in near-isogenic wheat lines susceptible and resistant to the RWA. Isoelectric focusing (IEF) and non-denaturing IEF gels revealed the existence of multiple isoenzymes of chitinase with a wide range of isoelectric points, ranging from 9.7 (basic) to 3.4 (acidic). Seven bands were observed in the resistant and susceptible untreated plants (pI values of approximately 9.5, 9.2, 8.8, 8.5, 8.4, 7.8 and 5.0) on IEF gels. RWA infestation resulted in the expression of an additional band (approximately pI 5.5) whereas ethylene exposure resulted in the induction of two additional isoenzymes (approximately pI 4.1 and 6.8). Mechanical wounding also induced the isoenzyme with pI of 4.1. Isoelectrical focusing (IEF) in liquid medium confirmed the multiple isoenzymes obtained with IEF gels. Five different peaks with chitinase activity were detected in control plants (pI values of 9.2, 7.8, 5.0, 4.1, and 3.6) after IEF. RWA infestation, ethylene and mechanical wounding caused the induction of additional isoenzyme peaks. Differential expression of chitinolytic activity was also observed between the near-isogenic wheat lines.
Keywords: Chitinase expression; Ethylene; Hydrolase; Isoelectric focusing; Isoenzymes; Mechanical wounding; Russian wheat aphid infestation.
Abbreviations: GlcNAc, b-1,4-linked 2-acetamino-2-deoxy-D-glucose; IEF, isoelectric focusing; IWF, intercellular washing fluid; NAD MDH, Malate dehydrogenease (NAD: EC 1.1.1.37); PR, pathogenesis related; RWA, Russian wheat aphid.
Introduction
Chitinase [poly{1,4-(N-acetyl-b-D-glucosaminide)} glycanohydrolase], (EC 3.2.1.14) catalyses the hydrolysis of chitin, a polymer of unbranched chains of b-1,4-linked 2-acetamido-2-deoxy-D-glucose (GlcNAc; N-acetylglucosamine) (Jeuniaux, 1966; Mauch and Staehelin, 1989). This enzymatic degradation of chitin to produce GlcNAc is performed by a chitinolytic system, which has been found in micro-organisms, plants, and animals (Flach et al., 1992). The chitinolytic enzymes are traditionally divided into two main classes: (1) endochitinases and (2) N-acetyl glucosaminidases (sometimes termed chitibiase, EC 3.2.1.30). The existence of a third class of enzyme, exochitinase, has been suggested (Robbins et al., 1988). Endochitinases randomly hydrolyse GlcNAc polymers, eventually giving diacetylchitobiose as
the major product. N-acetyl glucosaminidases preferentially act on a dimer (Gooday, 1990). The exochitinase also catalyses progressive release of diacetylchitobiose units from the non-reducing ends of chitin chains.
The physiological role of chitinases in the general metabolism of plant cells has not been documented. Indeed, a general role is difficult to envisage, because its substrate, chitin, does not occur in higher plants. It has been postulated that plants produce chitinase in order to protect themselves from chitin-containing parasites (Abeles et al., 1970; Bell, 1981; Boller, 1985) but, until recently, direct evidence supporting this hypothesis was lacking (Boller, 1985). However, in 1988, Roberts and Selitrennikoff reported that an endochitinase purified from barley, was capable of inhibiting the growth of Trichoderma reescei, Alternaria alternata, and Neurospora crassa. In addition, Mauch et al. (1988) reported that, in combination, chitinase and b-1,3-glucanase act synergistically to inhibit fungal growth. These results, therefore, support the hypothesis that the in vivo role of these pathogenesis-related proteins is to protect the host from invasion by fungal pathogens and that, as such, they are an integral component of a general disease resistance mechanism.
3Present address: Forestry and Agricultural Biotechnology Institute, University of Pretoria, Pretoria, 0001, South Africa.
4Corresponding author. Tel: +27-51-401-2776; Fax: +27-51-488- 772; E-mail: bothaam@fabi.up.ac.za