Bot. Bull. Acad. Sin. (1998) 39: 107_112
Huh et al. Allozyme variation of Pyrola
Allozyme variation and population structure of Pyrola japonica in Korea
Man Kyu Huh1, Su Dong Chung, and Hong Wook Huh
Department of Biology Education, Pusan National University, Pusan, 609-735, The Republic of Korea
(Received July 10, 1997; Accepted January 3, 1998)
Abstract. Enzyme electrophoresis was used to estimate genetic diversity and population structure of Pyrola japonica KLENZE in Korea. The percent of polymorphic loci within the enzymes was 58.8%. Genetic diversity at both the species level and at the population level was high (Hes = 0.226; Hep = 0.179, respectively) whereas the extent of the population divergence was relatively low (GST = 0.143). FIS, a measure of the deviation from random mating within the 14 populations, was 0.184. An indirect estimate of the number of migrants per generation (Nm = 1.50) indicates that gene flow is moderate among Korean populations of the species. In addition, analysis of fixation indices revealed a substantial heterozygosity deficiency in some populations and at some loci. This indicates that some populations sampled may have been substructured largely due to rhizotamous spread and restricted gene flow coupled with founder effects and genetic drift.
Keywords: Genetic diversity; Population structure; Pyrola japonica.
Introduction
Many perennial plants multiply by both sexual and asexual reproduction. Numerous plant species can produce offspring that are genetically identical to each other and to the maternal plant (Fryxell, 1957). This broad-sense asexual reproduction may be accomplished by vegetative spread or production of sexual propagules. Most plants, especially rhizomatous and stoloniferous species, have physical connections among ramets although their persistence is highly variable among species and habitats (Sobey and Barkhouse, 1977).
Studies of the genetic structure of apomictic plant populations have received revitalized interest in the past decade as a result of electrophoretic techniques, which allow us to better assess the genotypic composition of populations. A well-established general belief holds that asexually reproducing species lack genetic diversity and can be considered evolutionary "dead-ends." Various studies have shown that asexually reproducing plants can be much more genetically diverse than originally thought (Ellstrand and Roose, 1987). Clearly, descriptive genetic work on both sexual and asexual plant populations is needed as well. Despite the importance of information on genetic variation and population genetic structure for conservation purposes, detailed studies of the levels and distribution of genetic variation are not available for most species in Korea, for either sexually or asexually reproducing plants.
Pyrola japonica Klenze (Pyrolaceae), a widespread herb occurring throughout cooler, more temperate regions of the Northern Hemisphere is most abundant in the boreal and Arctic areas (Woodland, 1991). Leaves of this spe
cies are evergreen, alternate, and simple with stipules lacking. Flowers are regular, perfect, and hypogynous. In this species, we have compiled and quantitatively analyzed what is known of the genetic structure of populations to examine whether any trends occur both within and among populations.
The purpose of this study was: 1) to estimate how much total genetic diversity is maintained in the species; 2) to describe how genetic variation is distributed within and among populations; 3) to assess genetic structure of P. japonica; and 4) to characterize the population genetic structure to determine whether patches of the species are comprised of a single clone or a mixture of genets.
Materials and Methods
Sampling Procedure
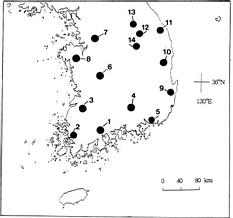
Pyrola japonica KLENZE was collected from fourteen natural populations in Korea (Figure 1). One leaf per plant was sampled during 1996 to 1997. More than 30 plants were collected from each population. Leaves gathered from natural populations were stored in plastic bags for 1_2 weeks in a refrigerator until electrophoresis was carried out.
Enzyme Electrophoresis
Leaves were homogenized by mechanical grinding to release enzymes from cell and organellar membranes with Tris-HCl grinding buffer-PVP solution described in Soltis et al. (1983). Electrophoresis was performed using 10% starch gel. Gel and electrode buffer systems and enzyme staining procedures from Soltis et al. (1983) were used to assay ten enzyme systems; peroxidase (PER), isocitrate dehydrogenase (IDH), glutamate oxaloacetate transami
1Corresponding author. Fax: (82-51) 514-8576; Tel: (82-51) 510-2698 or 510-2697.