Bot. Bull. Acad. Sin. (1998) 39: 113_118
Liu et al. Root formation and indole-3-acetic acid
Effect of hormone treatment on root formation and endogenous indole-3-acetic acid and polyamine levels of Glycine max cultivated in vitro
Zin-Huang Liu1, Wei-Chang Wang, and Yan-Sin Yen
Department of Biology, National Sun Yat-Sen University, Kaohsiung, Taiwan 804, Republic of China
(Received October 16, 1997; Accepted February 4, 1998)
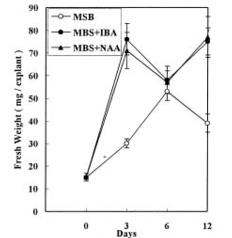
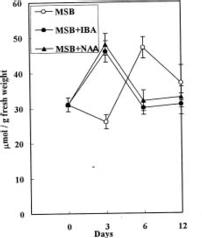
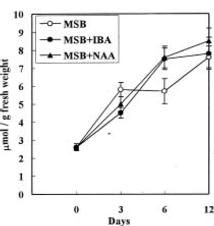
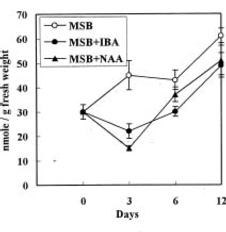
Abstract. Accumulation of endogenous indole-3-acetic acid (IAA) in soybean hypocotyl explants was found during adventitious root formation in naphthaleneacetic acid (NAA) and indole-3-butyric acid (IBA) treatments. The auxin-induced root formation was accompanied by increasing levels of putrescine. We suggest that the exogenously applied auxin (IBA and NAA) may act on polyamine synthase and IAA oxidase activity.
Keywords: Adventitious root; Glycine max; Indole-3-acetic acid; Polyamine.
Abbreviations: IAA, indole-3-acetic acid; IBA, indole-3-butyric acid; NAA, naphthaleneacetic acid.
Introduction
It is well known that root formation involves intensive mitotic activity and metabolic changes accompanied by changes of polyamine levels (Friedman et al., 1985; Jarvis et al., 1985; Burtin et al., 1990). Polyamines are considered important in cell division because they stimulate DNA synthesis (Kaur-Sawhney et al., 1980). Polyamine biosynthetic enzyme activity and polyamine levels increase before DNA replication (Cohen et al., 1984). Moreover, auxin seems to be an universal inducer of adventitious roots. Several reports have indicated a positive correlation between endogenous indole-3-acetic acid (IAA) levels in cuttings and the number of adventitious roots produced per cutting (Weigel et al., 1984; Alvarez et al., 1989). Past publications on the role of auxin in adventitious root induction were mainly based on experiments with exogenously applied synthetic auxin. However, there have been far fewer investigations into the effect of exogenously applied auxin on endogenous IAA and polyamine levels in plant tissue. In this study, we examine the effect of two synthetic auxins, naphthaleneacetic acid (NAA) and indole-3-butyric acid (IBA) on adventitious root induction using soybean hypocotyl explants.
Materials and Methods
Plant Tissue Culture
Soybean seeds (Glycine max) strain Tk5 were obtained from the Asia Vegetable Research and Development Center in Taiwan. Seeds were surface-sterilized for 1 min
in 70% ethanol and soaked in 1% sodium hypochlorite for 10 min. Seeds were rinsed three times in sterile distilled water. Seedlings were grown in darkness at 27°C, and relative humidity (RH) was maintained between 70 to 80%. Young seedlings (5-days old) about 5 cm in height were used for tissue culture. Five mm hypocotyl were cut and cultured on test medium. The basal medium (MSB medium) consisted of inorganic nutrients as found in Murashige-Skoog medium (Murashige and Skoog, 1962), and vitamins of Gamborg's B5 medium (Gamborg et al., 1968), plus 250 mg/L casein hydrolysate supplemented with 3% sucrose. Root formation without intermediate growth of callus was promoted by the addition of 2 mg/L indole-3-butyric acid (IBA) or 2 mg/L naphthaleneacetic acid (NAA) to the culture medium.
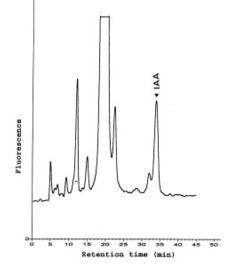
Analysis of Indole-3-Acetic Acid in Soybean Hypocotyl Explants
Soybean hypocotyl explants were cultured on three different media and grown in the incubator as described in darkness. Hypocotyl explants (adventitious roots were removed), were extracted after culture for certain days according to the experiments. Portions of explant tissue including the basal part that might be affected by the hormone treatment, usually 5 to 8 g fresh weight, were homogenized in 60 mL of 80% methanol containing sodium ascorbate and Butylated hydroxytoluene (BHT) (Dunlap and Guinn, 1989). The ground tissues were filtrated with suction through Whatman No.1 paper. Methanol in the filtrate was removed by rotary flash evaporation (RFE) at 35°C. The pH of the aqueous residues was adjusted to 8.0 and partitioned twice against an equal volume of ethyl acetate to remove phenolic compounds and other impurities. The pH of the aqueous fraction was adjusted to 2.5, then
1Corresponding author. Fax: 886-75253609; E-mail: zhliu@ mail.nsysu.edu.tw