Bot. Bull. Acad. Sin. (1998) 39: 119_123
Saha et al. Effect of time of nitrogen application on spikelet differentiation and degeneration of rice
Effect of time of nitrogen application on spikelet differentiation and degeneration of rice
Abhijit Saha1, R.K. Sarkar2, and Y. Yamagishi
Tsukuba International Agricultural Training Centre (TIATC), Tsukuba, Japan
(Received November 24, 1997; Accepted February 12, 1998)
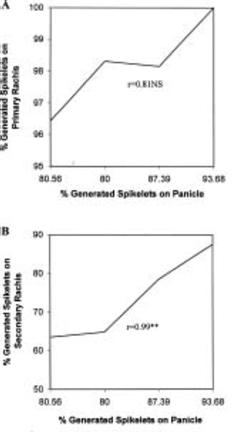
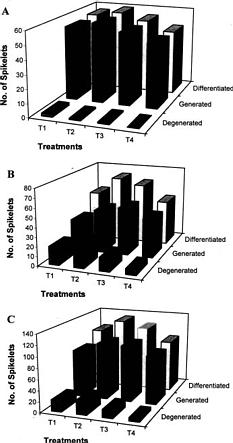
Abstract. A study was conducted in the field to identify the N application stages which greatly determine spikelet differentiation and degeneration of rice (Oryza sativa L.). Spikelet differentiation and degeneration were greatly influenced by time of N application. N application at the neck-node differentiation stage significantly increased spikelet differentiation and that at the active meiosis stage significantly reduced spikelet degeneration. N absorption at the spikelet differentiation and heading stages was significantly correlated with both greater spikelet differentiation and percent degeneration, respectively. Degeneration was mainly induced by the degeneration of spikelets on the secondary rachis-branch. Four equal splits of 100 kg N/ha at the early tillering, active tillering, neck-node differentiation, and active meiosis stages were found good for efficient rice production.
Keywords: Oryza sativa L.; Spikelet degeneration; Spikelet differentiation.
Abbreviations: AM, active meiosis; AT, active tillering; DAT, days after transplanting; NND, neck-node differentiation; RCB, randomized complete block; SPD, spikelet differentiation; SRD, secondary rachis-branch differentiation.
Introduction
Differentiation and degeneration of spikelets are the two important processes that determine final spikelets per panicle. In reproductive phase two stages, the SRD and AM stages, are the most critical in determining final bearing spikelets per panicle (Matsushima, 1992). At the SRD stage, spikelets are ready to be born, and their development is positively increased. At the AM stage, it is decreased by degeneration; thus, the final number of spikelets is determined by the difference between the number of grains differentiated at the SRD stage and the number of grains degenerated in the later period. Climatic, biological and nutritional status at those stages greatly influences the spikelet determination process and subsequently the yield.
Among nutritional factors, N plays a very important role in the differentiation and degeneration of spikelets (Wada and Cruz, 1990). Split application of N is very often used in rice cultivation to increase its availability in the critical growth stages. Therefore, this experiment was undertaken (1) to investigate influence of N spilts at different growth stages on the differentiation and degeneration of spikelets and (2) to identify the suitable time of N topdress for good harvest with reasonable panicles and spikelets.
Materials and Methods
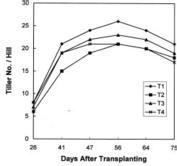
Twenty-two-day old rice (Oryza sativa L. cv. Kinuhikari) seedlings were transplanted in the experimental field at Tsuskuba International Agricultural Training Centre on May 10, 1995 at 30 × 18 cm2 spacing. The experiment was laid-out in loamy mixed mesic soil in RCB design with 3 replications. The treatments were application of total 100 kg N/ha in the form of urea (46% N) at different stages of rice growth as follows: T1-all at a time at 7 DAT ; T2-in three equal splits at 7 DAT, AT and NND stages; T3-in three splits at the ratio of 50:25:25 at 7 DAT, NND and SPD stages; T4-in three splits at the ratio of 50:25:25 at 7 DAT, SRD and AM stages. Active tillering was identified as the stage when the tillering rate per day was the highest. NND, SRD and SPD stages as mentioned by Yoshida (1981) were identified through microscopic observation and measurement of the young panicle length. The AM stage, mentioned by Matsushima (1992) and De Datta (1981), was identified as the stage when the collar of the flag leaf and that of the penultimate leaf coincided at the same level. Basal fertilizer of 120 kg P2 O5/ha and 80 kg K2 O/ha were also applied in all the plots one day before transplanting. During flowering 20 kg K2 O/ha was also top dressed. Other cultural management practices were adopted as and when necessary.
Differentiation and degeneration of spikelets on both primary and secondary rachis-branches were studied from the panicle samples collected after maturity. Differentiated spikelets were the total spikelets primordia born on the panicle, and generated and degenerated spikelets were
1Corresponding author. Present Address: Scientific Officer, BRRI Regional Station, Post Box 58, Comilla-3500, Bangladesh. Tel: 88-081-6945.
2Present address: Senior Scientist CRRI, Cuttack, Orissa, India.