Bot. Bull. Acad. Sin. (1998) 39: 147_152
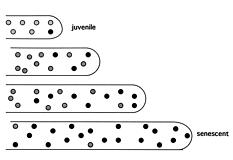
Griffiths Senescence plasmid
(Invited review paper)
The kalilo family of fungal plasmids
Anthony J. F. Griffiths
Department of Botany, University of British Columbia, Vancouver, Canada V6T 1Z4
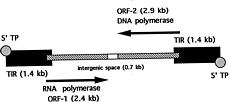
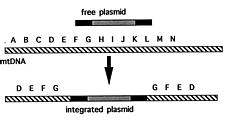
Abstract. A. J. F. Griffiths (1997). The prototypic kalilo plasmid is approximately 9 kb long and has the structure of an invertron. There are 5'-bound terminal proteins, a long terminal inverted repeat (TIR) and two ORFs corresponding to viral RNA and DNA polymerases. Kalilo plasmids have been found in several different heterothallic and one pseudohomothallic species of Neurospora from around the world. No examples have been found in true homothallic species. The prototypic plasmid was found in N. intermedia on the Hawaiian island of Kauai. It causes death by integration into the mtDNA, creating an inserted form flanked by long symmetrical repeats of the mtDNA flanking one side of the insertion point. The plasmid can be transmitted horizontally through heterokaryosis or sexually via the maternal parent. Spread is potentially explosive and can be prevented by cell incompatibility or by suppressor mutations. The related plasmid LA-kalilo has been found in N. tetrasperma from Louisiana, USA. This is essentially the same as kalilo but shows a short deletion in the middle of each TIR. This plasmid has been found not to insert into mtDNA. In a related genus Gelasinospora, also from Louisiana, a relative called Gel-kalilo has been found. This plasmid shows many differences from kalilo, but is still obviously homologous over large regions. This plasmid also does not integrate into the mtDNA. Lack of integration can be due to structural differences or to host genome suppression. The plasmids have arrived in their present hosts by a combination of vertical descent from a common ancestor and horizontal asexual transmission.
Keywords: Mitochondria; mtDNA; Neurospora; Plasmid, Senescence.
Contents
Introduction 147
The Kalilo Prototype 148
Discovery 148
Structure 148
Distribution 148
Transmission 149
Expression 149
Structural Variants 150
Suppressors 150
LA-Kalilo 150
Gel-Kalilo 151
Evolution 151
Literature Cited 152
Introduction
Plasmids are replicating DNA molecules that are additional to the normal genomic components. They contain few genes and so are dependent on the normal operation of the genomic machinery for their propagation and survival. In this sense, they are parasitic molecules, drawing on not only the information of the host but also on its energy reserves. Bacterial plasmids carry genes that seem to
benefit the host and are of adaptive value. Genes for resistance to environmental toxins are the best example of this. However, it must be said that little is known of the cost/benefit ratios or the evolutionary dynamics of bacterial plasmids in natural populations. Plasmids are not as commonly encountered in eukaryotic organisms. No plasmids have ever been found in animals, though several have been found in plants, and many in fungi (Griffiths, 1995). The reasons for this `kingdom specificity' are not known.
As in bacteria, fungal plasmids are either linear or circular. Linear plasmids are more common than circular plas
Fax: 604-822-9179; E-mail: agriff@unixg.ubc.ca