Bot. Bull. Acad. Sin. (1998) 39: 153_160
Yang et al. Phylogenetic position of Raphanus in relation to Brassica species
Phylogenetic position of Raphanus in relation to Brassica
species based on 5S rRNA spacer sequence data
Yau-Wen Yang1,3, Pey-Feng Tseng1, Pon-Yean Tai1 and Cheng-Ju Chang2
1Institute of Botany, Academia Sinica, Taipei, Taiwan, Republic of China
2Institute of Biomedical Science, Academia Sinica, Taiwan, Republic of China
(Received August 23, 1997; Accepted March 4, 1998)
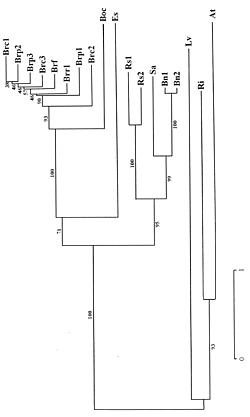
Abstract. Based on RFLP analysis, two evolutionary lineages for Brassica diploid species have been proposed. These are (I) the "nigra" lineage and (II) the "rapa/oleracea" lineage. The phylogenetic relationship of Raphanus species to these two lineages is still unclear because chloroplast and mitochondrial DNA genomic restriction site variation suggests that Raphanus is more closely related to the "rapa/oleracea" lineage, whereas nuclear RFLPs and other lines of evidence suggest that Raphanus belongs to the "nigra" lineage. Here, we present evidence of the intergenic spacer of nuclear 5S rRNA to support that Raphanus is more closely related to the "nigra" lineage than to the "rapa/oleracea" lineage. Genetic polymorphism within species is also discussed.
Keywords: Brassica; Phylogeny; Raphanus; 5S rRNA spacer.
Introduction
Brassica is one of the most important plant groups, containing species widely used in our daily life. Brassica rapa (n=10), B. nigra (n=8), and B. oleracea (n=9) are the three basic groups with three amphidiploid species, B. napus (n=19), B. juncea (n=18) and B. carinata (n=17) derived from interspecific hybridization between pairs of these diploid species, B. rapa × B. oleracea, B. rapa × B. nigra and B. nigra × B. oleracea, respectively (U, 1935). Based on chloroplast DNA (Warwick and Black, 1991), mitochondrial DNA (Palmer and Herbon, 1988), and nuclear DNA variation (Song et al., 1988; 1990) the phylogenetic relationships in Brassica and its related genera have been proposed. Brassica species can be divided into two evolutionary pathways: the "nigra" lineage and the "rapa/oleracea" lineage. Raphanus is thought to be closely related to the Brassica species. However, its relation to either lineage remains unsolved. Based on chloroplast and mitochondrial DNA restriction site variation (Warwick and Black, 1991; Palmer and Herbon, 1988), Raphanus was proposed to be more closely related to the "rapa/oleracea" lineage, but nuclear RFLPs and other RAPD data have suggested that Raphanus is more closely related to the "nigra" lineage (Song et al., 1990; Thormann et al., 1994).
5S rRNA genes are arranged in tandom arrays in the nuclear genome with highly conserved regions, separated by the non-transcribed intergenic spacer (IGS) that may vary in length or sequence between and even within spe
cies (Gerlach and Dyer, 1980; Long and Dawid, 1980). IGS evolves rapidly and is informative at the level of genus and species, so it has been chosen to study the phylogenetic relationships between and within genera (Appels et al., 1989, 1992; Baum and Appels, 1992; McIntyre et al., 1992; Moran et al., 1992; Playford et al., 1992; Reddy and Appels, 1989; Udovicic et al., 1995). Bhatia et al. (1993) demonstrated great polymorphism in IGS within B. rapa and related species. However, the phylogenetic relationship of Raphanus to these two lineages has not been studied using IGS sequences. Here we present data from the IGS of nuclear 5S rRNA to support the hypothesis that Raphanus is more closely related to the "nigra" lineage.
Materials and Methods
Plant Material
A total of twelve accessions of plant materials, which include seven accessions of B. rapa, one accession of B. oleracea, two accessions of R. sativus, one accession of Rorippa indica and one accession of Lepidium virginicum were used for PCR amplification of 5S rRNA sequences (Table 1). Among them, four accessions were provided by the Asian Vegetable Research and Development Center (AVRDC), and rest of them were collected by the authors. Also, another six 5S rRNA sequences from Genbank, which include sequences from B. rapa (X60723), Eruca sativa (X63524), B. nigra (X65710, X65711), Sinapis alba (X56866) and Arabidopsis athaliana (M65137) were used in this study (Bhatia et al., 1993; Campell et al., 1992; Capesius, 1991; 1993; Singh et al., 1994).
3Corresponding author.